Михаил Виленчик - Биологические основы старения и долголетия

- Название:Биологические основы старения и долголетия
- Автор:
- Жанр:
- Издательство:Знание
- Год:1987
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Виленчик - Биологические основы старения и долголетия краткое содержание
Биологические основы старения и долголетия - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
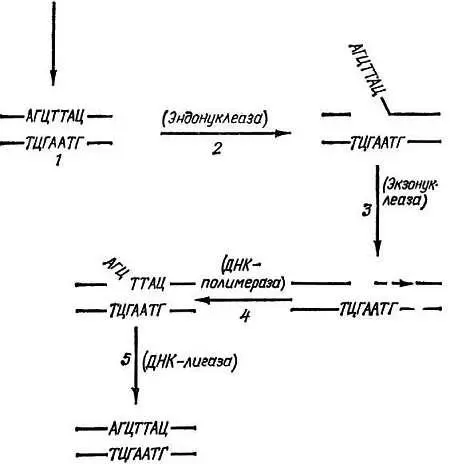
Рис. 6. Схема работы эксцизионных (вырезающих) механизмов репарации ДНК.
1.Индукция повреждения (реакция одного из оснований ДНК с ОН· или Н2О2 или другим эндогенным генотоксическим метаболитом; тепловое повреждение или попадание в ДНК квантов УФ, рентгеновского или другого вида ионизирующего излучения). 2. Узнавание повреждения и образование в ДНК разрыва. 3. Выщепление (вырезание) здорового участка ДНК, прилежащего к поврежденному. 4. Репаративный синтез ДНК. 5. Сшивание концов ДНК. В скобках указаны ферменты, катализирующие соответствующий этап репарации ДНК
Первый из них — "узнавание" повреждения в ДНК и образование вблизи него "надреза" (мы продолжаем проводить аналогию процесса репарации с работой хирурга). Такую операцию осуществляет фермент эндонуклеаза, разрушающий модифицированную нить ДНК. Затем подключается другой фермент — экзонуклеаза, который выщепляет модифицированное основание, а вместе с ним обычно и значительное количество "здоровых" оснований. В результате в ДНК образуется брешь. Поскольку же вторая нить остается целой, то, используя ее как матрицу, третий фермент — ДНК — полимераза заполняет эту брешь. Процесс репарации завершает четвертый фермент — лигаза. Этот фермент связывает вновь синтезированный участок ДНК с тем концом, который остался после процесса выщепления.
Рассмотренный пример наглядно показывает: в процессе репарации происходит синтез ДНК. Этот синтез связан не с делением клетки, а с восстановлением ДНК, поэтому его называют репаративным. Интенсивность его легко определить, добавляя в среду, где находятся клетки, меченый тимидин — вещество, которое входит в состав только ДНК. Теперь, чтобы ответить на вопрос о том, происходил ли "ремонт" ДНК, т. е. замещение отдельных ее оснований, достаточно установить, является ли ДНК клетки, не приступающей к делению, радиоактивной.
С помощью такого сравнительно простого приема нами и было получено доказательство того, что репарация ДНК происходит не только в том случае, когда клетки повреждаются физическими или химическими агентами, т. е. в необычных условиях, но и в таких условиях, которые можно считать нормальными, физиологическими. Поэтому такой процесс получил название спонтанного репаративного синтеза ДНК.
Кстати, вывод, который неизбежно следует из факта протекания спонтанного репаративного синтеза, состоит в том, что ДНК клетки в процессе нормального метаболизма повреждается. Следовательно, биологическая стабильность генома, его надежность обеспечиваются и "волшебной палочкой квантовой химии", т. е. стабильностью ее химических связей (Э. Шредингер), и системой репарации ДНК, функционирующей в клетке.
Только что рассмотренную систему репарации ДНК (по причинам, теперь, наверное, понятным читателю) называют вырезающей или эксцизионной (от англ. exision — вырезать). Одна из принципиальных особенностей репаративного синтеза ДНК, протекающего при такой репарации, состоит в том, что он осуществляется в различные фазы клеточного цикла. "Обычный" же синтез ДНК, в результате которого происходит удвоение количества ДНК и поэтому называемый редупликативным (обычно говорят "репликативный"), протекает лишь в определенную фазу жизни клетки. Таким образом, редупликативный синтез ДНК является запрограммированным, протекающим в S-фазу клеточного цикла, а репаративный — незапрограммированным, или внеплановым.
Система эксцизионной репарации ДНК и механизм ее редупликации резко отличаются по чувствительности к оксимочевине. Репаративный синтез не ингибируется под влиянием концентраций этого вещества в дозах, при которых редупликативный синтез ингибируется почти полностью. Такое различие было найдено не только в культуре клеток, но и в тканях взрослых крыс и мышей. В мозге этих животных митотическая активность глиальных клеток резко уменьшается, а нейроны взрослых животных вообще лишены способности делиться. Поэтому мы полагали, что интенсивность синтеза, связанного с делением глиальных клеток, и внепланового синтеза — величины одного порядка. Но если это так, то внеплановый синтез ДНК в мозге взрослых животных можно было определить по его резистентности к оксимочевине. Таким образом, суть проведенного нами вместе с Т. М. Третьяк эксперимента состояла в том, что животным вводили оксимочевину с таким расчетом, чтобы в течение нескольких часов примерно на 95 % ингибировать редупликативный синтез ДНК (концентрации были подобраны на основании исследования синтеза ДНК в печени подопытных крыс, а также в модельной системе на культивируемых фибробластах человека). Сравнение интенсивности редупликативного синтеза в клетках головного мозга крыс первых дней жизни и взрослых животных заставляет предположить, что наряду с репарацией спонтанных повреждений ДНК в них протекает, вероятно, и другой процесс, также не связанный с делением клеток.
Наиболее вероятное объяснение полученных нами данных состоит в том, что включение меченого тимидина в ДНК мозга взрослых крыс в значительной степени определяется репаративным (внеплановым) синтезом ДНК, который индуцируется спонтанными генетическими повреждениями. Это заключение подкрепляется результатами исследования включения 3Н-тимидина в ДНК "нейрональных" и "глиальных" фракций ядер двухлетних крыс. Было найдено, что удельная активность ДНК нейрональных ядер в расчете на 1 мкг ДНК лишь в 2 раза меньше удельной активности ДНК ядер глиальных клеток. Включение тимидина в ДНК глиальных ядер определяется в значительной степени тем, что в части глиальных ядер протекает и редупликативный синтез ДНК. Ведь известно, что деление глиальных клеток в мозге крыс и других млекопитающих происходит в течение всей жизни. Что же касается включения тимидина в нейрональные ядра, то оно, очевидно, определяется в основном репаративным синтезом ДНК.
Однако при исследовании динамики содержания меченого тимидина в ДНК головного мозга крыс были получены данные, которые невозможно объяснить лишь исходя из предположения, что внеплановый синтез ДНК связан только с репарацией спонтанных повреждений ДНК.
Эти данные показывают, что наряду с репаративным синтезом ДНК, необходимым для устранения спонтанных повреждений ДНК, в головном мозге протекают и другие формы "внепланового" синтеза ДНК. Одна из этих форм может включать обратную транскрипцию, т. е. процесс синтеза ДНК на матрице РНК, катализируемый обратной транскриптазой. Другая форма синтеза может состоять в перестройке определенных генов клеток головного мозга крыс, аналогичной перестройке иммуноглобулиновых генов В-лимфоцита при его дифференцировке. В этом процессе может участвовать часть тех ферментов, которые катализируют процесс репарации спонтанных повреждений ДНК.
Читать дальшеИнтервал:
Закладка:









