Инна Шустрова - КОШКИ. ГЕНЕТИКА И ПЛЕМЕННОЕ РАЗВЕДЕНИЕ

- Название:КОШКИ. ГЕНЕТИКА И ПЛЕМЕННОЕ РАЗВЕДЕНИЕ
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Инна Шустрова - КОШКИ. ГЕНЕТИКА И ПЛЕМЕННОЕ РАЗВЕДЕНИЕ краткое содержание
КОШКИ. ГЕНЕТИКА И ПЛЕМЕННОЕ РАЗВЕДЕНИЕ - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
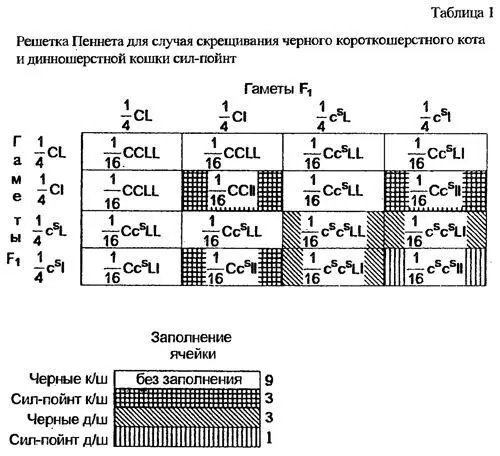
Третий закон Менделя звучит так: аллели каждого гена распределяются в потомстве независимо от аллелей другого гена.
Почему это происходит? Дважды гетерозиготные особи первого поколения могут образовывать четыре типа половых клеток: CL Cl csL csl, а объединяются эти гаметы при оплодотворении равновероятным и случайным образом. Чтобы было проще рассчитать варианты, возникающие при таком объединений, пользуются так называемой решеткой, или таблицей Пеннета, в которой по горизонтали откладываются все варианты гамет одного из родителей, а по вертикали - другого (в рассматриваемом случае гаметы родителей одинаковых типов). На пересечении в каждой клетке записывается образующийся при данной комбинации половых клеток генотип потомка, а на его основе уже определяется фенотип (табл.1).
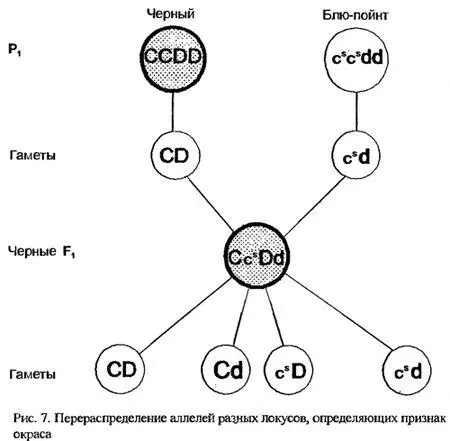
А что получится, если скрестить того же черного кота с кошкой голубого сиамского окраса (блю-пойнт)? Вроде бы обе характеристики относятся к одному и тому же признаку - окрасу. В первом поколении, понятно, все потомки останутся черными. А вот при скрещивании их между собой, кроме ожидаемых черных и блю-пойнтов, появятся и чисто-голубые, и сил-пойнты. При этом распределение окрасов у этих котят будет соответствовать третьему закону Менделя. Получается, что голубой окрас наследуется независимо от сиамского? Это действительно так. Голубой - осветленный - окрас возникает в результате действия рецессивного аллеля d из совсем другого локуса - Dilution, никакого отношения к локусу Colour не имеющего. Аллель d в гомозиготной форме вызывает осветление любого окраса - черного в голубой, красного в кремовый, сил-пойнта- в блю-пойнт...
Доминантный аллель D обеспечивает интенсивный окрас. Так что генотипы родительских особей следует записывать как CC DD (для кота) и cscs dd (для кошки), а их потомков - как двойных гетерозигот: Ccs Dd (рис.7)
Соответственно может быть составлена и решетка Пеннета (табл.2).
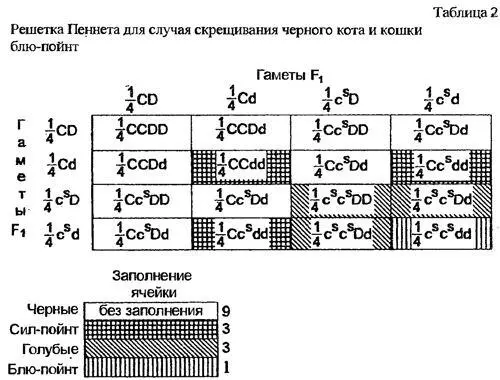
Стало быть, такой "признак", как окрас, в реальности определяется не геном одного локуса, а многими генами, рассматривать которые можно только в парных сочетаниях (белый - цветной, пегий - без белых пятен, интенсивный - осветленный).
Однако, если бы вся наследственность подчинялась только менделевским законам, вся племенная работа свелась бы к элемент тарным арифметическим подсчетам. Как говорят генетики, Менделю в: свое время очень повезло, что он случайно начал работу со столь; просто наследуемыми генами. Уже упоминавшееся ограничение двух; из этих законов — то, что они справедливы только при больших выборках.
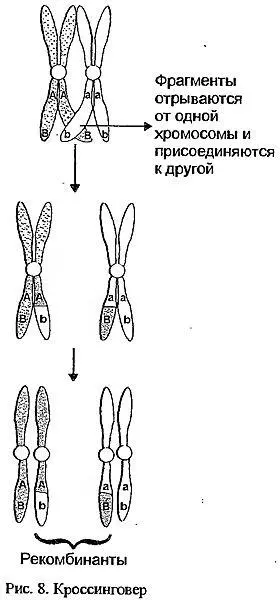
Признание генетиками менделевских законов и попытки применить их к различным организмам показали, что существуют и более значительные ограничения. Причем первым было обнаружено отклонение от третьего закона — так называемое сцепление генов. В самом деле, если при дигибридном скрещивании гены, по котором гомозиготны родители, находятся на разных парных хромосомах, в процессе перераспределения (формирования половых клеток и оплодотворении) эти Пары хромосом как бы независимы друг от друга. Ну а если оба гена расположены на одной хромосоме? Их аллели будут наследоваться вместе, то есть сцеплено - и никакого "независимого наследования" признаков в потомстве второго поколения не будет, а будут наблюдаться исключительно или преобладать только родительские комбинации признаков. Причем чем ближе друг к другу расположены локусы таких генов, тем меньше потомков будет нести отклонения от родительских пар признаков. Казалось бы, таких "отклоняющихся" потомков вообще не должно быть. Однако в процессе деления при образовании половых клеток есть стадия, на которой гомологичные хромосомы обмениваются участками. Чем больше расстояние на хромосоме между двумя локусами генов, тем больше вероятность, что между ними произойдет такой обмен – кроссинговер(рис.8).
Гены, находящиеся на значительном удалении друг от друга, вообще ведут себя как не сцепленные, и наоборот, тесно сцепленные гены обычно наследуются как единое целое. Основываясь на частоте появления в потомстве "отклоняющихся" особей, можно рассчитать, насколько далеко друг от друга расположены на хромосоме те или иные гены. Таким образом составляют так называемые генетические карты разных видов животных. У кошек к настоящему времени мало изучены группы сцепления генов (во всяком случае, тех, которые в первую очередь интересуют заводчиков). Однако, работая с новыми мутациями, список которых постоянно пополняется, рано или поздно фелинолог может столкнуться с этим явлением.
Другой случай отклонения от законов Менделя тоже связан со сцеплением: так называемым сцеплением с полом.
Попробуем скрестить черного кота с красной кошкой.(Разумеется, как и в предыдущих случаях, будем считать, что оба родителя гомозиготны по интересующему нас гену красного окраса, не принимая в расчет другие гены; фактически альтернативной парой признаков будет: красный - не красный (условно - черный)).
Никакого единообразия в первом поколении не наблюдается: все котики - красные, все кошечки - черепаховые, черно-красные. Если поставить обратное (реципрокное) скрещивание: красный кот и черная кошка, то все котята-самцы будут черные, а самки - опять-таки черепаховые (рис.9).
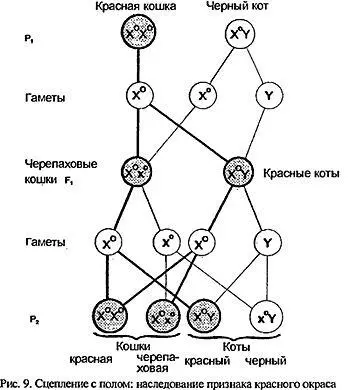
Дело в том, что локус Orange, доминантный аллель которого Оопределяет красный окрас, расположен на половой хромосоме X. А как мы уже говорили, две Х-хромосомы имеются только у самок; (геномный набор 18 пар аутосом + XX), тогда как у самцов Х- хромосома только одна, а вместо второй присутствует Y- хромосома(18 пар + XY). Y- хромосома по сравнению с Xочень мала, и большая часть локусов, характерных для X, на ней отсутствует. Однако именно Y определяет мужской путь развития организма. В процессах деления при образовании половых клеток X и Y ведут себя как гомологи, хотя реально таковыми не являются. В результате такой неполной гомологии между ними у самок образуется "избыток" генетического материала по сравнению с самцами. Чтобы такую несправедливость компенсировать, одна из самочьих X -хромосом переходит в неактивное состояние - принимает такую форму, что генетическая информация с нее не считывается. Которая именно из двух Х- хромосом в каждой конкретной клетке зародыша и в какой именно период перейдет в нерабочее состояние - заранее неизвестно, это процесс случайный. (Во всяком случае так считалось до недавнего времени. В последние годы появились работы, показывающие неслучайность инактивации Х- хромосом, или их избирательную, зависящую от генотипа самки, инактивацию.)
Читать дальшеИнтервал:
Закладка:





![Альма Либрем - Ведьмина генетика [СИ]](/books/1060227/alma-librem-vedmina-genetika-si.webp)
