Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Все это возможно в плотной водной среде, когда требуется обеспечить сквозной ток воды через жабры. В разреженной воздушной среде такой насос будет крайне неэффективным, ведь помимо разной плотности среды у дыхательного насоса и задачи другие. Нужно не гнать воздух сквозным потоком, а вентилировать слепой воздушный мешок. Но эту задачу первые тетраподы (земноводные) толком так и не решили. Это удалось лишь рептилиям. А у земноводных принцип нагнетания воздуха в легкие примерно тот же, что и у дышащих воздухом рыб. Они используют преобразованный аппарат жаберных дуг для расширения глоточной полости и, соответственно, для вдоха. Отработанный воздух из легких выдыхается за счет сокращения мускулатуры легких (а у двоякодышащих рыб он выходит за счет более высокого давления в воде по сравнению с воздушной поверхностью над водой, куда рыба выставляет голову для вдоха-выдоха).
Особенности жаберного аппарата, приспособленного для вдоха-выдоха за счет движения глотки, а не жаберной полости, хорошо отражаются в скелете и современных, и ископаемых девонских лопастеперых. Для более эффективного выведения углекислого газа из организма лопастеперые используют жабры, а земноводные, утратившие жабры, используют кожу («кожное дыхание»). Это создает массу ограничений, из-за которых земноводные так и не смогли далеко уйти от воды.
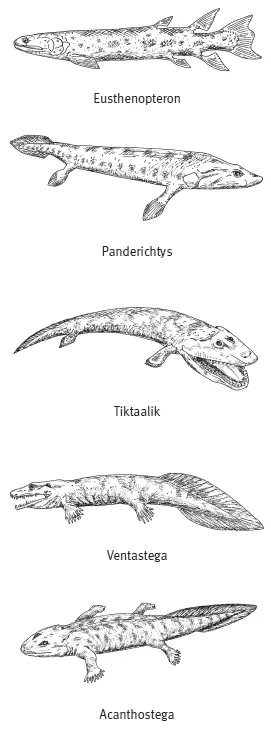
Но в воздушной среде жаберная крышка все равно не работала в качестве воздушного насоса, поэтому от нее пришлось отказаться, и это открыло наземным обитателям дополнительные возможности — косточки жаберной крышки ушли внутрь черепа. И начали формировать среднее ухо. Жаберная крышка частично редуцировалсь у тиктаалика. Это означает, что тот способ дыхания, которым продолжали пользоваться другие рыбы, стал ему больше не нужен. Тиктаалик дышал в основном воздухом при помощи легких и, возможно, кожей.
Раз у тиктаалика редуцировалась жаберная крышка, это означает, что ему пришлось научиться глотать без использования всасывающего жаберного насоса — рыбы втягивают частицы пищи с током воды. Тиктаалик глотал только за счет движения глотки и головы. У акантостеги, наследницы тиктаалика, еще сохранялись внутренние жабры, а у ихтиостеги они уже редуцировались (наружные жабры у личинок амфибий сохраняются и сегодня). Следовательно, последователи тиктаалика тоже глотали за счет движений головы и челюстей. В дальнейшем по мере редукции остатков жаберного аппарата у древних амфибий происходило постепенное развитие шейного отдела.
Таким образом, в строении дыхательной и кровеносной систем при переходе от рыб к тетраподам не было резких скачков, а большинство важных изменений произошло еще на «рыбьем» этапе. Каждое новшество в дыхании и кровообращении не создавалось на пустом месте, а лишь улучшало требуемые функции на основе развития имеющихся приспособлений.
—————
Плавники превращаются в пальцы: взгляд генетика
Недавно было обнаружено семейство генов, необходимых для развития плавников у рыб и получивших название actinodin ( and ). У наземных позвоночных этих генов нет. Если отключить у рыбы гены and , то, во-первых, в парных плавниках не развиваются эластичные плавниковые лучи — лепидотрихии (но в хвостовом плавнике они развиваются нормально), во-вторых — меняется характер работы ряда ключевых генов-регуляторов (в том числе hoxd13a и sonic hedgehog ), которые есть не только у рыб, но и у четвероногих и от которых зависит передне-задняя полярность зачатка конечности (будь то плавник или нога). Похожие изменения в работе этих генов-регуляторов у четвероногих, возникающие в результате некоторых мутаций, приводят к полидактилии (многопалости).
По-видимому, потеря генов and древними четвероногими привела к утрате лепидотрихий в парных конечностях, однако в хвостовом плавнике лепидотрихии могли сохраниться. Именно так обстоит дело у древнейших четвероногих, обнаруженных в ископаемой летописи, таких как акантостега и ихтиостега. У этих животных уже не было плавниковых лучей на парных конечностях, но они еще сохранялись в плавниковой складке, окаймлявшей хвост. У более ранних переходных форм, таких как тиктаалик, плавниковые лучи на парных конечностях имелись.
Логично предположить, что утрата генов and и вызванная ею дестабилизация работы регуляторных генов у древнейших четвероногих создала предпосылки для развития у них большого числа (до восьми) почти одинаковых пальцев. В дальнейшем у четвероногих восстановился исходный, «нормальный» характер работы генов-регуляторов, причем теперь их работа уже не зависела от утраченных генов and . Результатом этого могло стать автоматическое сокращение числа пальцев до пяти. Так изучение генетики современных животных помогает понять молекулярные механизмы событий, произошедших сотни миллионов лет назад ( Zhang et al., 2010 ).
—————
Мы рассмотрели ряд переходных форм, связывающих лопастеперых рыб с примитивными четвероногими. Совершенно непонятно, где в этом ряду находится искомый великий «макропереход». Приобретение выростов пищевода, в которые можно набирать воздух? Нет, это просто адаптация рыб к плохо аэрируемым водоемам. Аналогичные органы воздушного дыхания (например, в виде модификаций ротовой полости) возникали и в более поздние времена у разных рыб, оказавшихся в схожих условиях. Приобретение костей поясов конечностей у эустеноптерона? Едва ли, ведь это тоже была чисто «рыбья» адаптация к большей подвижности плавников. Может, назначить этот рубеж по приобретению подвижного сустава у лап, т. е. где-то между эустеноптероном и тиктааликом? Но ведь это просто адаптация рыб к ползанию по дну мелких луж и переползанию из одного пересохшего водоема в другой, развивавшаяся к тому же долго и постепенно. Еще есть вариант — присвоить «макро»-титул тиктаалику, у которого передние конечности отделились от черепа и стали свободными лапками… хотя и это всего лишь улучшенный вариант адаптации для передвижения по дну на мелководье. Утрата плавниковых лучей и приобретение пальцев? При большом желании можно, конечно, объявить эту небольшую модификацию дистальных участков конечностей тем самым эпохальным «превращением рыбы в амфибию». Действительно, именно в этом месте непрерывного ряда переходных форм специалисты проводят формальную грань между рипидистиями и тетраподами. Но не лучше ли честно признать, что палеонтологические данные показывают просто длинный ряд постепенных маленьких изменений, каждое из которых само по себе никак не тянет на «макрособытие»?
Интервал:
Закладка:








