Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Стохастическая изменчивость обычно настолько мала, что совсем не бросается в глаза и часто игнорируется биологами. Особенно ничтожна она у животных: у растений стохастическая изменчивость проявляется сильнее, как показывает пример с сиренью. Подавление стохастической изменчивости происходит из-за того, что реальные генетические программы развития буквально до отказа набиты стабилизирующими регуляторными контурами, основанными на обратных связях. Адаптивный смысл этих генно-регуляторных сетей как раз в том и состоит, чтобы свести элемент случайности к минимуму и сделать развитие максимально стабильным (помехоустойчивым) [94].
Попытки спроектировать какое-нибудь интересное существо в программе EvoDevo поначалу всегда приводят к нестабильному результату. Первый успешный набор правил (генотип) создает нужное нам существо не всегда, а лишь изредка. При этом даже лучший из вариантов фенотипа, как правило, будет далек от того идеала, к которому мы стремились, а остальные и вовсе получатся уродцами. Чтобы улучшить результат, т. е. уменьшить стохастичность онтогенеза и стабилизировать нужный вариант фенотипа, приходится вносить в генотип стабилизирующие правила, основанные на отрицательных обратных связях и дополнительных регуляторах. Эти дополнения на первый взгляд могут показаться «избыточными». Но без них не обойтись. Например, если клетки при таких-то условиях начинают выделять морфоген, даже не рассчитывайте, что его количество будет одним и тем же всякий раз при запуске развития зиготы. Если для нормального развития необходимо, чтобы количество морфогена было постоянным, придется добавить в генотип специальные стабилизирующие правила, примерно такие: если концентрация морфогена А больше 0,5, уменьшить производство морфогена А на 10 %. Это и есть отрицательная обратная связь: если чего-то стало слишком много, нужно принять меры, чтобы его стало меньше. Увеличение показателя должно стимулировать его уменьшение. Иначе все пойдет вкривь и вкось.
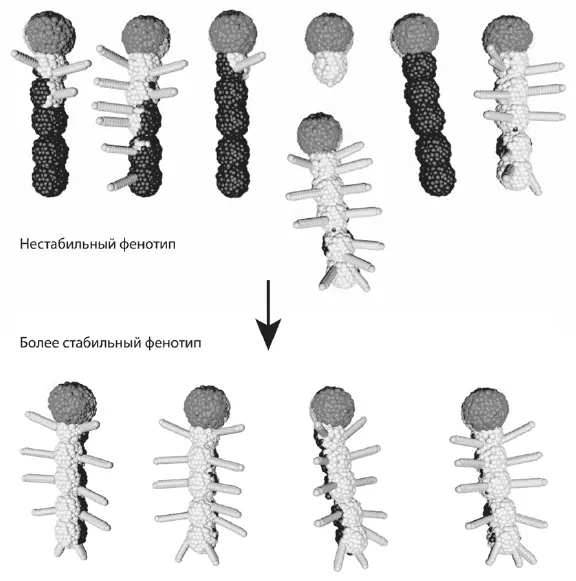
Пример «онтогенетической стохастики» в программе EvoDevo . Генотипы, содержащие недостаточное количество стабилизирующих обратных связей, дают нестабильный результат. При этом желаемый («хорошо приспособленный») фенотип будет получаться лишь изредка. Чтобы перейти от нестабильного фенотипа (вверху) к более стабильному (внизу), нам пришлось усложнить генотип (программу развития) на 50 %.
Чтобы стабилизировать развитие «правильного» признака — в модели это признак, который нам нравится, а в природе это признак, повышающий приспособленность, — приходится усложнять программу развития. Например, чтобы перейти от нестабильного фенотипа, показанного на рисунке сверху, к более стабильному, показанному внизу, нам пришлось увеличить количество правил, составляющих генотип, на 50 %. В программе EvoDevo это делаем мы, пользователи. В природе этим занимается дарвиновский эволюционный механизм, основанный на мутациях и отборе. Адаптивное преимущество получают те генотипы, которые производят оптимальный (хорошо приспособленный) фенотип с наибольшей вероятностью и с наименьшей зависимостью от всякого рода случайностей — внешних или внутренних. Генотип, обеспечивающий развитие оптимального фенотипа лишь при температуре от 20 до 21 °C, проиграет в конкуренции генотипу, генерирующему такой же фенотип в более широком диапазоне температур (если только эволюция идет не в термостате). Вместо температуры можно сюда подставить любые факторы внешней или внутренней среды и вообще любые «помехи», в том числе связанные с изначально присущей онтогенезу стохастикой, о которой говорилось выше.
Введение дополнительного правила в генотип модельного существа аналогично появлению в ходе эволюции нового сайта связывания ТФ, ставящего активность одного из генов — регуляторов развития в зависимость от какого-нибудь нового фактора. Например, от концентрации морфогена, который ранее не влиял на этот ген. Это все равно что добавить дополнительное условие в оператор условного перехода (команду «если… то…») в компьютерной программе.
—————
Избыточность и обратные связи в работе шпемановского организатора
Главная функция организатора Шпемана состоит в создании стабильной дорзовентральной (спинно-брюшной) разметки эмбриона. Там, где концентрация активных белков из группы BMP (bone morphogenetic proteins) минимальна, клетки наружного слоя зародыша превращаются в нервные. Формируется нервная пластинка — зачаток центральной нервной системы. Там будет спинная сторона. Нервная пластинка в свою очередь индуцирует развитие других органов и структур. Остальные клетки наружного слоя под действием морфогенов BMP — а их больше на будущей брюшной стороне зародыша — становятся клетками эпителия. Спинной (шпемановский) организатор производит белок-морфоген хордин, который блокирует действие BMP. Там, где много хордина, мало активных молекул BMP, в результате в этой области зародыша формируется спина.
Казалось бы, вот и хорошо: система из двух взаимодействующих морфогенов, BMP и chordin, вполне достаточна для дорзовентральной разметки зародыша. Какие-то дополнительные ухищрения здесь, наверное, не нужны. Не тут-то было!
Как выяснилось, помимо основного регуляторного взаимодействия «хордин блокирует BMP» в создании дорзовентральной полярности у позвоночных участвует целый ряд морфогенов и связей между ними. Спинной организатор «общается» с брюшным организующим центром посредством белков-морфогенов, выделяемых в межклеточное пространство. Все эти морфогены тем или иным способом влияют на дорзовентральный градиент BMP, усиливая или ослабляя его. Отрицательные обратные связи, которыми изобилует эта регуляторная сеть, делают градиент BMP устойчивым к разнообразным помехам и нарушениям. Например, если производство BMP по какой-то причине увеличивается, это приводит к росту экспресии белка Sizzled, который подавляет активность белка Tolloid, который в свою очередь выводит из строя хордин, инактивирующий BMP (см. рисунок ниже). Получается типичный регуляторный контур, основанный на отрицательной обратной связи: больше BMP → больше Sizzled → меньше Tolloid → больше Chordin → меньше BMP .
Благодаря наличию нескольких таких контуров градиент BMP оказывается на редкость устойчивым. Генно-инженерные эксперименты показали, что для того, чтобы серьезно нарушить этот градиент — и получить, например, полностью «дорзализованный» эмбрион, у которого весь наружный слой клеток превращается в нервную ткань, или полностью «вентрализованный» эмбрион вообще без нервной пластинки, — недостаточно вывести из строя какой-то ключевой ген-регулятор. Даже без хордина, даже без BMP4 дорзовентральный градиент все равно худо-бедно формируется за счет дополнительных регуляторных контуров и морфогенов-«дублеров». Нужно выключить сразу 3–4 гена, чтобы нарушения развития стали действительно катастрофическими ( De Robertis, 2009 ). Вот что значит настоящая помехоустойчивость!
Читать дальшеИнтервал:
Закладка:








