Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Разумеется, естественный отбор не стал бы создавать столь поразительный запас прочности только на случай прихода любопытных биологов, которые начнут отключать в эмбрионах гены. В природе и без нас хватает неприятных неожиданностей: перемены климата и химического состава среды, мутации, неудачные комбинации родительских генов… Все это может нарушить работу тех или иных генных систем. Поэтому в общем случае генотипы, обеспечивающие более помехоустойчивое развитие, получают преимущество над генотипами, срабатывающими только в тепличных условиях. Ну а если жизнь становится более предсказуемой — как, например, у внутриклеточных паразитов — может произойти редукция стабилизирующих систем и упрощение генетической программы развития.
—————
Изменения регуляторных участков генов — регуляторов развития играют важнейшую роль в эволюции многоклеточных. С несколькими примерами мы познакомились в главе 5. Но до сих пор мы исходили из негласной презумпции однозначного соответствия между генотипом и фенотипом : изменился геном — изменился признак.
Пора внести важное уточнение. Мы должны помнить, что само это соответствие не с неба падает. По умолчанию оно вовсе не однозначно, а для того, чтобы оно стало таковым, необходимы специальные стабилизирующие адаптации, которые должны постепенно развиваться в ходе эволюции.
Это согласуется с идеей о творческой роли стабилизирующего отбора, которую развивал великий советский биолог И. И. Шмальгаузен (1884–1963). Отбор на стабильность должен вести к усложнению программы развития. Отбор, просто отсеивающий уродцев (отклонения от «нормы», т. е. от оптимального в данных условиях фенотипа), в перспективе способствует развитию новых генно-регуляторных контуров, повышающих стабильность воспроизведения «нормального» фенотипа.
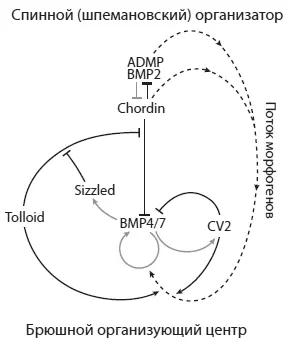
Регуляторная сеть, обеспечивающая дорзовентральную (спиннобрюшную) разметку эмбриона шпорцевой лягушки Xenopus . Из De Robertis, 2009 .
Генотип определяет не фенотип как таковой, а норму реакции, т. е. диапазон возможных фенотипов, которые могут быть реализованы при данном генотипе. «Выбор» конкретного фенотипа часто зависит от условий, в которых происходит развитие. Но даже генетически идентичные особи, развивающиеся в одинаковых условиях, все равно будут хоть немного, но разными. И чем хуже у них работают стабилизирующие генно-регуляторные контуры, тем сильнее будет случайная вариабельность.
Наличие стохастической составляющей в генетической программе развития ярко проявляется в неполной пенетрантностимногих мутаций. Суть этого явления, замеченного еще в 1925 году выдающимися генетиками Н. В. Тимофеевым-Ресовским (1900–1981) и Д. Д. Ромашовым (1899–1963), состоит в том, что одна и та же мутация у одних особей проявляется в фенотипе, а у других нет. Это зависит от условий среды и «генетического контекста», т. е. от других генов в геноме (что неудивительно), но даже с поправкой на оба эти фактора многие мутации все равно имеют нестабильный фенотипический эффект.
О молекулярных механизмах неполной пенетрантности известно немного. Их изучение осложняется тем, что генные сети, управляющие развитием, обычно крайне громоздки и на первый взгляд избыточны. Входящие в их состав гены связаны сложнейшей сетью взаимодействий, содержащей многочисленные петли положительных и отрицательных обратных связей, так что трудно понять, где именно в этой паутине возникает непредсказуемость.
Идеальный объект для таких исследований — наш старый знакомец червь Caenorhabditis elegans , у которого генетическая регуляция развития предельно упрощена, как, впрочем, и само развитие [95].
Биологам из Массачусетского технологического института и Принстонского университета (США) удалось выяснить причины неполной пенетрантности нескольких мутаций гена skn-1 ( Raj et al., 2010 ). Эти мутации приводят (или не приводят, так как пенетрантность неполная) к тому, что потомки бластомера(зародышевой клетки) E , которые должны стать клетками кишечника, остаются недифференцированными, и кишечник у эмбриона не формируется.
Роль главного «переключателя» в развитии кишечника играет ген elt-2 . Он активирует сотни других генов, необходимых для превращения бластомеров в кишечные клетки. Чтобы сам elt-2 своевременно включился, требуется работа генов end-3 и end-1 . Они в свою очередь активируются геном skn-1 , причем активация осуществляется как напрямую, так и опосредованно, через ген med-1/2 .
Известно несколько мутаций гена skn-1 , нарушающих развитие кишечных клеток. Все эти мутации в гомозиготном состоянии летальны, и в этом смысле пенетрантность у них стопроцентная. Однако у некоторых мутантных эмбрионов перед смертью формируется нормальный кишечник, а у других — нет. В этом отношении пенетрантность неполная.
Чтобы выяснить причины неполной пенетрантности, авторы воспользовались новым методом флюоресцентной окраски молекул РНК, который они сами и изобрели. Метод позволяет увидеть и даже подсчитать молекулы матричной РНК (мРНК), считанные с определенного гена. Каждая отдельная молекула мРНК становится видна как светящаяся точка. Это дает уникальную возможность определять уровень активности гена в индивидуальных клетках.
Оказалось, что у эмбрионов с мутантным геном skn-1 в клетках, которые должны стать кишечником, med-1/2 не включается вовсе, а end-3 работает очень слабо. Что касается гена end-1 , то его активность становится непредсказуемой. В норме этот ген на определенной стадии развития эмбриона начинает работать в клетках, которые должны стать кишечником, причем уровень его активности во всех этих клетках примерно одинаков. У мутантных эмбрионов end-1 начинает работать в разных клетках с разной силой, а в некоторых не включается вовсе, так что общая картина получается хаотичной.
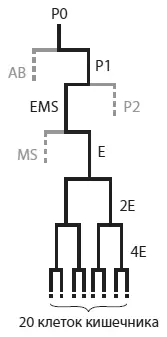
«Родословная» клеток кишечника в развитии эмбриона C. elegans . Кишечник взрослого червя состоит из 20 клеток. Все они являются потомками бластомера E, который происходит от бластомера EMS, а тот — от P1. Бластомер P1 образуется в результате первого деления оплодотворенной яйцеклетки (P0). «Братом» бластомера EMS является бластомер P2 — организатор, формирующий передне-заднюю полярность зародыша при помощи морфогена Wnt. Из Raj et al., 2010 .
Этот хаос на следующем этапе регуляторного каскада приводит к одному из двух альтернативных результатов: либо ген elt-2 включается, и тогда клетки превращаются в кишечник, либо elt-2 остается выключенным, и тогда кишечник не формируется. Как выяснилось, выбор одного из двух исходов зависит от того, достигнет ли активность end-1 определенного порогового уровня. Однако и тут сохраняется доля неопределенности. Если активность end-1 ниже пороговой, elt-2 точно не включится и кишечника не будет. Если же активность end-1 выше пороговой, то это еще не гарантирует включения elt-2: он либо включится, либо нет. Раз включившись, elt-2 уже будет работать нормально сколько ему положено, потому что этот ген активирует сам себя.
Читать дальшеИнтервал:
Закладка:








