Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Чтобы окончательно убедиться в том, что активность wingless является достаточным условием для появления пятен у D. guttifera , авторы провели еще одну серию генно-инженерных экспериментов. На этот раз в эмбрионы D. guttifera впрыскивали ген wingless , взятый у D. melanogaster и соединенный с регуляторным участком, заимствованным у дрожжей. Смысл заключался в том, чтобы получить трансгенных мух D. guttifera , у которых области экспрессии wingless в крыльях были бы расположены как-то иначе. В конце концов такие мухи были получены: у них возникла новая область экспрессии wingless вдоль одной из продольных жилок. В полном соответствии с теоретическими ожиданиями у этих мух появилась и новая темная полоса на крыльях, идущая вдоль этой жилки.
Авторы также проанализировали пигментацию крыльев, экспрессию wingless и наличие или отсутствие регуляторного элемента vs у других видов дрозофил. В итоге удалось восстановить ход эволюции узора на крыльях.
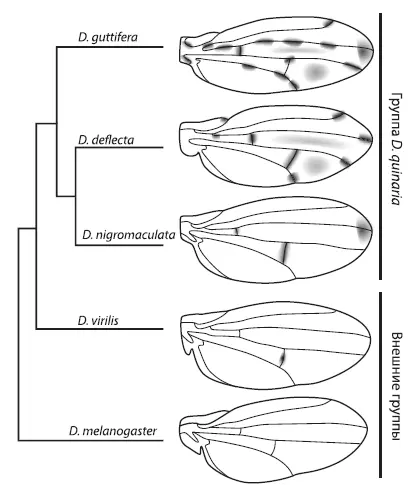
Слева — эволюционное дерево, показывающее родственные отношения между видами дрозофил с различными узорами на крыльях. Самый сложный орнамент — у D. guttifera . Справа — схема эволюции узора. Из Werner et al., 2010 .
Исходно у дрозофил ген wingless экспрессировался вдоль поперечных жилок, но не работал у концов продольных жилок и в колоколовидных сенсиллах. В гене yellow не было регуляторного элемента vs , способного реагировать на wingless , а крылья не имели орнамента. Это исходное состояние сохранилось у D. melanogaster . Затем в одной из групп дрозофил появился элемент vs, что привело к формированию связи между экспрессией wingless и пятнами на крыльях [69]. Первые темные пятна поэтому появились там, где исходно экспрессировался wingless , — вдоль поперечных жилок.
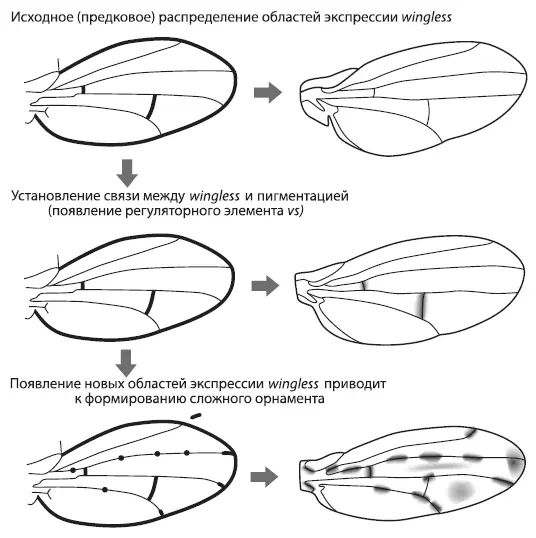
После того как пигментация стала зависеть от wingless , у эволюции появилась возможность создавать новые узоры, меняя экспрессию этого гена. Ген wingless имеет сложную и пока еще слабо изученную систему регуляции, но в целом его работа ориентируется на базовую разметку и ключевые «опорные точки» развивающегося организма. Эта привязанность wingless к опорным точкам сохранилась и в ходе дальнейших изменений его экспрессии в крыльях. У некоторых видов возникли новые области экспрессии wingless на концах продольных жилок, и там появились пятна. Только у одного вида — D. guttifera — орнамент крыльев усложнился еще сильнее за счет появления дополнительных областей экспрессии wingless в шести колоколовидных сенсиллах.
Получается, что замысловатый орнамент крыльев D. guttifera развился на основе существовавшего ранее плана строения крыла. Возможно, примерно таким же образом, отталкиваясь от опорных точек морфогенетической разметки крыла, сформировались узоры на крыльях и у других насекомых, включая бабочек. Но, чтобы это доказать, придется еще немало потрудиться.
Здесь мы видим, как изменение двух регуляторных участков одного гена (гена yellow ) привело к формированию сложного признака. Изменение это было таково, что направило уже существующий регуляторный каскад по новому маршруту: старые персонажи в новой пьесе.
—————
Как трехкамерное сердце стало четырехкамерным
Выход позвоночных на сушу был связан с развитием легочного дыхания, что потребовало радикальной перестройки кровеносной системы. У дышащих жабрами рыб один круг кровообращения, а сердце, соответственно, двухкамерное (состоит из одного предсердия и одного желудочка). У наземных позвоночных — трех- или четырехкамерное сердце и два круга кровообращения. Один из них (малый) прогоняет кровь через легкие, где она насыщается кислородом. Затем кровь возвращается к сердцу и попадает в левое предсердие. Большой круг направляет обогащенную кислородом (артериальную) кровь ко всем прочим органам, где она отдает кислород и по венам возвращается к сердцу, попадая в правое предсердие.
У животных с трехкамерным сердцем кровь из обоих предсердий попадает в единый желудочек, откуда она затем направляется и к легким, и ко всем прочим органам. При этом артериальная кровь смешивается с венозной. У животных с четырехкамерным сердцем в ходе развития изначально единый желудочек подразделяется перегородкой на левую и правую половины. В результате два круга кровообращения оказываются полностью разделены: бедная кислородом кровь попадает из правого предсердия в правый желудочек и идет оттуда к легким, насыщенная кислородом — из левого предсердия только в левый желудочек и идет оттуда ко всем прочим органам.
Формирование четырехкамерного сердца было необходимой предпосылкой развития теплокровности у млекопитающих и птиц. Ткани теплокровных потребляют очень много кислорода, поэтому им необходима «чистая» артериальная кровь, максимально насыщенная кислородом. А смешанной артериально-венозной кровью могут довольствоваться холоднокровные позвоночные с трехкамерным сердцем. Трехкамерное сердце характерно для амфибий и большинства рептилий, хотя у последних намечается частичное разделение желудочка на две части (развивается неполная внутри-желудочковая перегородка). Настоящее четырехкамерное сердце развилось независимо в трех эволюционных линиях: у крокодилов, птиц и млекопитающих. Это яркий пример параллельной эволюции.
Биологам из США, Канады и Японии удалось частично расшифровать молекулярно-генетические основы этого важнейшего эволюционного события ( Koshiba-Takeuchi et al., 2009 ). Ключевую роль в нем сыграли изменения в работе гена Tbx5 . Этот ген, кодирующий регуляторный белок, по-разному экспрессируется в развивающемся сердце у амфибий (шпорцевой лягушки Xenopus ) и теплокровных (курицы и мыши). У первых он равномерно экспрессируется по всему будущему желудочку, у вторых его экспрессия максимальна в левой части зачатка (в будущем левом желудочке) и минимальна справа. А как обстоит дело у рептилий?
Выяснилось, что у рептилий — ящерицы и черепахи — на ранних эмбриональных стадиях ген Tbx5 экспрессируется так же, как у лягушки, т. е. равномерно по всему будущему желудочку. У ящерицы все так и остается до конца развития. Как и у лягушки, у ящерицы не формируется ничего похожего на перегородку (хотя бы частичную) между желудочками.
Что касается черепахи, то у нее на поздних стадиях формируется градиент экспрессии — такой же, как у цыпленка, только выраженный слабее. Иными словами, в правой части желудочка активность гена постепенно снижается, а в левой остается высокой. Таким образом, по характеру экспрессии Tbx5 черепаха занимает промежуточное положение между ящерицей и курицей. То же самое можно сказать и о строении сердца. У черепахи формируется неполная перегородка между желудочками, но на более поздних стадиях, чем у цыпленка. Сердце черепахи занимает промежуточное положение между типичным трехкамерным (как у амфибий и ящериц) и четырехкамерным, как у крокодилов и теплокровных.
Читать дальшеИнтервал:
Закладка:








