Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
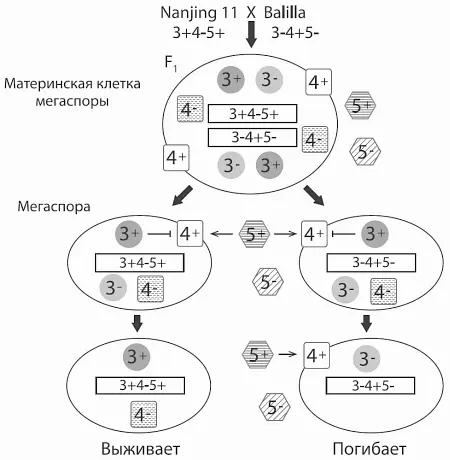
Схема, объясняющая пониженную плодовитость гибридного потомства от скрещивания сортов Nanjing 11 (indica, генотип 3+4−5+) и Balilla (japonica, генотип 3−4+5−). Материнская клетка (диплоидная) дает в результате мейоза четыре гаплоидные мегаспоры. Материнская клетка, как и все гибридное растение, гетерозиготна по трем рассматриваемым генам и поэтому содержит как рабочие (3+, 4+, 5+) , так и нерабочие (3−, 4−, 5−) варианты трех белков. В результате мейоза из нее образуются гаплоидные мегаспоры двух типов: с генотипом indica и с генотипом japonica (генотипы не перекомбинируются, потому что все три гена расположены вплотную друг к другу на хромосоме). Поначалу в мегаспорах сохраняются все шесть белков, унаследованных от материнской клетки, но впоследствии в них остаются только белки, закодированные в геноме самой мегаспоры. При этом белок 5+ , выделяющийся в межклеточное пространство, действует на все мегаспоры независимо от того, закодирован ли он в их геноме. Мегаспоры с генотипом japonica погибают, потому что на них действуют совместно оба белка-убийцы 5+ и 4+ , а белка-защитника 3+ у них нет. Мегаспоры с генотипом indica выживают, потому что у них нет убийцы 4+ и есть защитник. Из Yang et al., 2012 .
Что касается сортов, которые свободно скрещиваются и с indica , и с japonica , то у них могут быть разные генотипы, например, 3−4−5− или 3+4+5−. Несовместимость проявляется, только когда с носителем аллеля 5+ скрещивается растение с генами 3− и 4+. Тогда у половины мегаспор не окажется защитника (3+) от действующих совместно «убийц» 5+ и 4+.
Для диких родственников культурного риса, в том числе для предкового вида Oryza rufipogon , характерен генотип 3+4+5+, т. е. активны и оба «убийцы», и «защитник». Скорее всего, такой же генотип был и у первых одомашненных сортов риса. Вероятно, у диких предков все три гена зачем-то нужны (хотя мы пока не знаем зачем), а переход к «культурной» жизни на возделываемых полях сделал их излишними. Впоследствии у двух групп сортов, indica и japonica , закрепились мутации, выводящие из строя некоторые гены этого комплекса. Внутри каждой группы мутации были взаимно совместимыми, потому что несовместимые комбинации, такие как 3−4+5+, отсеивались отбором. Но отбор не проверял мутации, закрепившиеся в одной группе, на совместимость с мутациями, закрепившимися в другой. Так и возникла частичная постзиготическая несовместимость между сортами — в полном согласии с моделью Добжанского — Мёллера.
—————
Ранние этапы видообразования могут сопровождаться геномными перестройками
Крупные геномные перестройки, такие как разрыв или слияние хромосом, могут привести к быстрому развитию постзиготической изоляции. Часто особи с разным числом хромосом либо вовсе не могут произвести потомство либо это потомство будет бесплодным. У гибридов первого поколения, получивших от отца и матери разное количество хромосом, в ходе мейоза хромосомы не могут должным образом разбиться на пары и распределиться по половым клеткам; в результате гибриды оказываются бесплодными.
Но это, конечно, не абсолютное правило. Будь оно абсолютным, все мутации, меняющие число хромосом, немедленно отсеивались бы отбором, а мы знаем, что это не так. Различия в числе хромосом не всегда препятствуют их нормальному спариванию в ходе мейоза. Хромосомы просто соединяются гомологичными участками: например, две половинки разорвавшейся хромосомы могут пристроиться к соответствующим половинкам своего неразорванного прототипа. И тогда гибридная особь произведет жизнеспособные половые клетки.
Есть виды, у которых число хромосом варьирует, а особи с разным числом хромосом сохраняют генетическую совместимость. Впрочем, такая ситуация вряд ли может сохраняться долго: плодовитость гибридов, скорее всего, будет все-таки понижена, поэтому отбор будет способствовать выработке дополнительных изолирующих механизмов. В итоге родительский вид разделится на два дочерних, различающихся по числу хромосом. Или измененный кариотип(хромосомный набор) будет отсеян отбором, и вид сохранит исходное число хромосом.
Межвидовая изменчивость по числу хромосом встречается гораздо чаще, чем внутривидовая. Например, у человека 23 пары хромосом, а у шимпанзе и других человекообразных — 24. Вторая хромосома человека возникла в результате слияния двух предковых хромосом, причем в месте слияния остался «шрам», который ни с чем не спутаешь: две расположенные «голова к голове» последовательности, типичные для концевых участков хромосом (теломер).
Если не учитывать многочисленные случаи полиплоидии, то самая большая вариабельность по числу хромосом среди видов одного рода наблюдается у бабочек-голубянок Agrodiaetus . В пределах этого рода диплоидное число хромосом варьирует от 20 до 268, хотя род появился и начал дивергировать недавно: 2,5–3,8 млн лет назад ( Lukhtanov et al., 2005 ) и все его виды очень похожи друг на друга. У позвоночных изменчивость тоже может быть значительной: например, у сомиков Corydoras диплоидное число хромосом колеблется от 44 до 102, у южноамериканских грызунов туко-туко ( Ctenomys ) — от 10 до 70.
Каким образом у недавно разделившихся видов формируются такие сильные различия по числу хромосом, если внутривидовая хромосомная изменчивость — явление довольно редкое? Возможны два сценария.
Во-первых, различия могут накапливаться постепенно, причем каждая перестройка (слияние или разрыв хромосом) соответствует одному акту видообразования. Такое «пошаговое» формирование межвидовых хромосомных различий, по-видимому, широко распространено в природе. Его начальные этапы можно наблюдать у некоторых видов с варьирующим числом хромосом, таких как домовая мышь и обыкновенная бурозубка.
Но возможен и другой сценарий: кратковременный взрывообразный рост хромосомной изменчивости внутри вида, за которым следует быстрое разделение исходного вида на несколько дочерних. Этот второй сценарий недавно удалось подтвердить В. А. Лухтанову из Зоологического института в Санкт-Петербурге и его испанским коллегам ( Lukhtanov et al., 2011 ). Исследователи описали удивительный случай высокого хромосомного полиморфизма в пределах одного вида бабочек, горошковой беляночки Leptidea sinapis . Этот вид встречается от Португалии и Испании на западе до Сибири на востоке.
Оказалось, что число хромосом у бабочек этого вида убывает с запада на восток — от 2n = 106 в испанских популяциях до 2n = 56 в Восточном Казахстане. Кроме того, число хромосом может варьировать в одной и той же популяции (такое наблюдается в Италии, Румынии и Казахстане).
Читать дальшеИнтервал:
Закладка:








