Елена Наймарк - Перспективы отбора

- Название:Перспективы отбора
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:2019
- Город:Москва
- ISBN:978-5-17-114115-8
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Елена Наймарк - Перспективы отбора краткое содержание
Перспективы отбора - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Ранее было показано, что на протяжении первых 20 000 поколений приспособленность росла с замедлением. Что будет дальше? Следует ли ожидать выхода на плато, то есть достижения постоянного уровня приспособленности, или же плато не получится и приспособленность будет продолжать понемногу расти? Этот вопрос – достижим ли вообще предел приспособленности – очень важен для биологов-теоретиков, ведь речь, по сути, идет о границах творческих возможностей эволюции.
Для начала ученые сравнили приспособленность бактерий из поколений № 40 000 и 50 000. Оказалось, что в среднем подопытные популяции за период смены 10 000 поколений повысили свою приспособленность на целых 3 %. Таким образом, даже после 40 000 поколений адаптация не прекратилась: микробы все еще продолжают накапливать полезные мутации и повышать свою приспособленность.
Затем исследователи проследили траекторию роста приспособленности, заставляя замороженных на разных этапах эксперимента микробов соревноваться с предками (рис. 1.1).
Статистический анализ полученных данных показал, что изменения приспособленности соответствуют степенной модели с неограниченным (хотя и замедляющимся) ростом. Это значит, что приспособленность вовсе не собирается выходить на плато. Иными словами, бактерии, по всей видимости, не намерены останавливаться на достигнутом. Несмотря на долгую жизнь в одних и тех же условиях, у бактерий все еще время от времени закрепляются новые мутации, в данных условиях полезные (напомним, что в эволюционной биологии слово “полезный” обычно используется в смысле “повышающий приспособленность”).
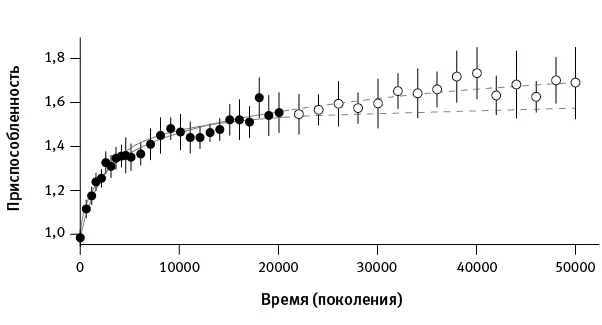
рис. 1.1.Рост приспособленности бактерий за 50 000 поколений. Точками (первые 20 000 поколений) и кружочками (следующие 30 000 поколений) показаны усредненные значения по всем подопытным популяциям. По вертикальной оси – относительная приспособленность (скорость роста по сравнению с предковым штаммом). Пунктиром показаны предсказания моделей: гиперболической (приспособленность асимптотически стремится к предельному уровню; нижняя кривая ) и степенной (неограниченный, хотя и замедляющийся рост приспособленности). Параметры моделей основаны на данных по первым 20 000 поколений. Видно, что гиперболическая модель хуже предсказывает динамику приспособленности за последующие 30 000 поколений, чем степенная. По рисунку из Wiser et al., 2013 .
Удивительная неисчерпаемость эволюционных возможностей, впервые столь наглядно продемонстрированная в эксперименте, имеет огромное значение для понимания эволюции. Ведь можно было предположить (многие так и думали), что в постоянных условиях, тем более в монокультуре, возможности для адаптивной эволюции быстро исчерпаются и система придет в состояние стазиса. В таком случае объяснить продолжающуюся эволюцию жизни на нашей планете можно было бы только непостоянством среды (абиотической и биотической).
По-видимому, запаса потенциально полезных мутаций хватит не только на 50 000 поколений бактерий, но и на 50 000 поколений наблюдающих за ними исследователей. Как ни странно, идущий безостановочно эволюционный процесс не приведет к какому-то невероятному, запредельному росту приспособленности. Ученые рассчитали, что если приспособленность будет и впредь увеличиваться по той же траектории (описываемой степенной моделью), то пятидесятитысячный по счету преемник Ленски увидит бактерий, удваивающих свою численность каждые 23 минуты. Это высокая скорость размножения для бактерий, но не какая-то неслыханная (численность бактерий предкового штамма удваивается за 55 минут).
Откуда же берутся новые полезные мутации? Ведь за время эксперимента каждая возможная нуклеотидная замена уже наверняка успела произойти хотя бы у одной бактерии! Основных причин, по-видимому, две. Первая связана с эпистазом – своего рода взаимодействием между мутациями. Например, мутация Б становится полезной только после того, как бактерия уже обзавелась мутацией А . Здесь нужно помнить еще и о том, что многие полезные мутации имеют негативные побочные эффекты. Мутация А может улучшить работу одной молекулярной системы, немного повредив другой. Допустим, первый эффект сильнее второго, поэтому в суммарном зачете мутация А оказывается “полезной” и поддерживается отбором. Но теперь, когда в популяции закрепилась мутация А , полезной окажется любая компенсаторная мутация Б , сглаживающая негативный побочный эффект мутации А .
Вторая причина неисчерпаемости запаса полезных мутаций состоит в том, что мутации с очень слабым полезным эффектом требуют очень долгого времени для закрепления в популяции. Такая мутация, появившись, с большой вероятностью будет вскоре потеряна из-за генетического дрейфа – случайных колебаний частот аллелей. Чтобы отбор начал хоть немного “помогать” такой мутации распространяться, число ее носителей должно стать довольно большим, а до тех пор она будет находиться целиком под властью безжалостного, неразборчивого дрейфа (см. Исследование № 4). Расчеты показывают, что подавляющее большинство вновь возникающих полезных мутаций теряются. Чтобы данная слабополезная мутация в конце концов все-таки закрепилась, она должна многократно появиться у разных бактерий независимо. Ленски и его коллеги рассчитали, что мутация, повышающая приспособленность бактерий на одну миллионную, должна возникнуть примерно 250 000 раз, прежде чем она наконец распространится и зафиксируется в популяции. Если принять во внимание частоту мутирования подопытных бактерий (в среднем одна мутация на 10 10 нуклеотидов за поколение) и размер их популяций ( эффективная численность каждой из них, с учетом ежедневных бутылочных горлышек , составляет примерно 33 000 000 клеток), получится, что такой мутации понадобится 100 000 000 поколений, чтобы “спастись” от дрейфа, и еще миллионы поколений, чтобы зафиксироваться (достичь стопроцентной частоты). Таким образом, накопление слабополезных мутаций в эксперименте Ленски вряд ли закончится в обозримом (и даже необозримом) будущем.
Причины замедления роста приспособленности связаны с тем, что каждая закрепившаяся полезная мутация делает последующие генетические усовершенствования в среднем менее полезными. Авторы называют этот эффект “эпистазом убывающей доходности” (по аналогии с экономическим законом убывающей доходности). Иными словами, сначала происходят наиболее радикальные адаптивные изменения, а затем идет все более тонкая настройка и оптимизация фенотипа .
Читать дальшеИнтервал:
Закладка:



![Екатерина Верхова - Иллюзия отбора [СИ]](/books/1060394/ekaterina-verhova-illyuziya-otbora-si.webp)