Дэвид Хьюбел - Глаз, мозг, зрение

- Название:Глаз, мозг, зрение
- Автор:
- Жанр:
- Издательство:Мир
- Год:1990
- Город:Москва
- ISBN:5-03-001254-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Дэвид Хьюбел - Глаз, мозг, зрение краткое содержание
В книге известного американского нейрофизиолога, лауреата Нобелевской премии, обобщены современные представления о том, как устроены нейронные структуры зрительной системы, включая кору головного мозга, и как они перерабатывают зрительную информацию. При высоком научном уровне изложения книга написана простым, ясным языком, прекрасно иллюстрирована. Она может служить учебным пособием по физиологии зрения и зрительного восприятия.
Для студентов биологических и медицинских вузов, нейрофизиологов, офтальмологов, психологов, специалистов по вычислительной технике и искусственному интеллекту.
Глаз, мозг, зрение - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
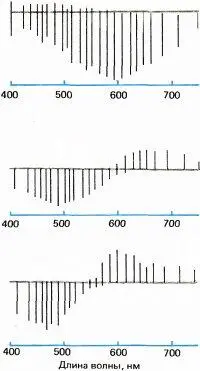
Рис. 125. Гуннар Светихин и Эдуард Мак-Никол регистрировали реакции горизонтальных клеток на цвет у костистой рыбы. Отклонения вниз от серой линии соответствуют гиперполяризации, а отклонения вверх — деполяризации.
В 1958 году Рассел де Валуа и его сотрудники зарегистрировали ответы, поразительно сходные со светихинскими, от клеток наружного коленчатого тела макака. Ранее с помощью поведенческих тестов де Валуа показал, что цветовое зрение макака и человека почти одинаково; например, соотношение, в котором нужно смешать два цветных луча, чтобы получить третий цвет, почти идентично у обоих видов. Поэтому можно думать, что макаки и люди обладают сходными механизмами на нижних уровнях зрительной системы, и мы, по-видимому, вправе сравнивать психофизику цвета у человека с физиологией у макака. Де Валуа обнаружил, что многие клетки коленчатого тела активировались рассеянным монохроматическим светом в диапазоне от одного конца спектра до точки перекреста, где реакция отсутствовала, и подавлялись светом во втором диапазоне — от точки перекреста до другого конца спектра. И вновь аналогия с цветовыми процессами Геринга была полной: де Валуа выявил оппонентно-цветовые клетки двух типов, красно-зеленые и желто-синие; у каждого типа смешение двух световых волн, длины которых на волновой шкале располагались симметрично относительно точки перекреста, приводило к взаимному погашению реакций точно так же, как в восприятии добавление синего к желтому или зеленого к красному порождает белый. Результаты де Валуа особенно хорошо согласовались с формулировками Геринга, поскольку две группы цветовых клеток имели максимумы реакции и точки перекреста точно в тех местах спектральной шкалы, чтобы одна группа могла отражать «желто-синие» свойства падающего света, а другая группа — «красно-зеленые».

Рис. 126. В типичном рецептивном поле типа 1 центр посылает возбуждающие сигналы от красных колбочек, а периферия — тормозящие сигналы от зеленых колбочек.
Следующий шаг состоял в том, чтобы рассмотреть рецептивные поля этих клеток, используя вместо рассеянного света маленькие цветные пятнышки; это мы и сделали в 1966 году совместно с Торстеном Визелом. У рецептивных полей большинства оппонентно-цветовых клеток де Валуа обнаружилась удивительная организация, которая до сих пор ставит нас в тупик. Клетки, как и у кошек по данным Куффлера, имели поля, разделенные на два антагонистических участка — центр и периферию; центр мог быть типа on или off. В типичном случае центр поля представлен исключительно красными, а тормозящая периферия — исключительно зелеными колбочками. Поэтому при красном свете как маленькое, так и большое пятно вызывает энергичную реакцию, поскольку центр избирательно чувствителен к длинноволновому свету, а периферия на него почти не реагирует; при коротковолновом свете маленькие пятна дают лишь очень слабую реакцию или не вызывают ее вовсе, а большие порождают сильное торможение с off-реакциями. При белом свете, содержащем короткие и длинные волны, малые пятна вызывают on-реакции, а большие не вызывают ответа.
Хотя нашим первым впечатлением было, что такая клетка из центра должна получать входные сигналы от красных, а с периферии — от зеленых колбочек, теперь кажется вероятным, что полное рецептивное поле включает две перекрывающиеся системы, как это показано на рис. 127. Как красные, так и зеленые колбочки распределены по довольно широкой круговой области, причем их количество максимально в центре и убывает по мере удаления от него. В центре красные колбочки сильно преобладают, но к периферии число их падает значительно быстрее числа зеленых колбочек. Поэтому небольшое пятнышко, вспыхивающее в центре и содержащее длинноволновую компоненту, будет весьма мощным стимулом для красной системы; даже если оно стимулирует и зеленые колбочки, их число по сравнению с общим числом связанных с клеткой зеленых колбочек будет слишком мало, чтобы составить какую-либо конкуренцию красной системе. Те же соображения приложимы и к клеткам с центром и периферией, описанным в главе 3, рецептивные поля которых тоже должны состоять из двух оппонентных круговых перекрывающихся зон с разными формами кривых зависимости чувствительности от координаты. Таким образом, периферия, вероятно, имеет не кольцеобразную, как думали вначале, а круговую форму. В отношении этих оппонентно-цветовых клеток обезьян предполагается (правда, без достаточных данных), что периферические области отражают вклад горизонтальных клеток.
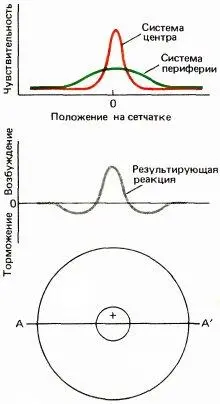
Рис. 127. Графики вверху отражают зависимость чувствительности нейрона (измеренной, например, по реакции на неподвижное очень маленькое световое пятнышко) от положения стимула на сетчатке вдоль линии AA, проходящей через центр рецептивного поля. Для клетки с r-центром и g –-периферией красное пятнышко дает островершинную кривую, а зеленое — значительно более широкую кривую. Нижний график отображает реакцию на белый или желтый свет, стимулирующий обе оппонентные системы, так что происходит вычитание вкладов двух систем. При этом красные колбочки доминируют в центре, что приводит здесь к on-реакциям, а зеленые — на периферии, что приводит к off-реакциям.
Реакция на диффузный свет — в данном случае on-реакции на красный, off-реакции на синий и зеленый и отсутствие реакции на белый свет — ясно показывают, что такая клетка должна регистрировать информацию о цвете. Но реакции на определенного рода белые края и отсутствие реакций на диффузный свет убеждают в том, что эта клетка связана также и с восприятием формы. Мы называем эти цветооппонентные клетки с центром и периферией клетками «типа 1».
Наружное коленчатое тело обезьяны, если вспомнить сведения из главы 4, содержит шесть слоев, причем четыре верхних слоя содержат множество мелких клеток, а два нижних — меньшее число крупных клеток. Мы находим клетки описанного выше типа 1 в верхних, или мелкоклеточных, слоях. Они различаются по типу колбочек, входящих в их центральную и периферийную системы, и по характеру центра, который может быть возбуждающим или тормозящим. Пример, представленный на рис. 126, мы можем обозначить как r +g –(с красным центром и зеленой периферией). Среди подтипов клеток, получающих входы от красных и зеленых колбочек, мы находим все четыре возможных варианта: r +g –, r –g +, g +r –, g –r +. Вторая группа клеток получает входы от синих колбочек из центра поля и от комбинации красных и зеленых (или, быть может, только от зеленых) — с периферии. Мы называем такие клетки «сине-желтыми», причем слово «желтые» здесь употреблено для краткости вместо «красные плюс зеленые».
Читать дальшеИнтервал:
Закладка:







![Дэвид Доу - Почини свой мозг. Программа восстановления нейрофункций после инсульта и других серьезных заболеваний [litres]](/books/1143582/devid-dou-pochini-svoj-mozg-programma-vosstanovlen.webp)

