Дэвид Хьюбел - Глаз, мозг, зрение

- Название:Глаз, мозг, зрение
- Автор:
- Жанр:
- Издательство:Мир
- Год:1990
- Город:Москва
- ISBN:5-03-001254-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Дэвид Хьюбел - Глаз, мозг, зрение краткое содержание
В книге известного американского нейрофизиолога, лауреата Нобелевской премии, обобщены современные представления о том, как устроены нейронные структуры зрительной системы, включая кору головного мозга, и как они перерабатывают зрительную информацию. При высоком научном уровне изложения книга написана простым, ясным языком, прекрасно иллюстрирована. Она может служить учебным пособием по физиологии зрения и зрительного восприятия.
Для студентов биологических и медицинских вузов, нейрофизиологов, офтальмологов, психологов, специалистов по вычислительной технике и искусственному интеллекту.
Глаз, мозг, зрение - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Неожиданно в 1978 году все изменилось. Нейроанатом из Сиэтла Маргарет Уонг-Райли обнаружила, что при окрашивании коры на фермент цитохромоксидазу в верхних слоях выявляется никогда не наблюдавшаяся ранее неоднородность — периодические темноокрашенные участки шириной около четверти миллиметра, разделенные промежутками около полумиллиметра. Цитохромоксидазу — участвующий в метаболизме фермент — содержат все клетки, и никто не мог даже подумать, что гистохимическая реакция на этот фермент позволит увидеть в коре что-либо интересное. Когда Уонг-Райли прислала нам свои микрофотографии, Торстен Визел и я заподозрили, что мы видим колонки глазодоминантности в поперечном разрезе и что большинство монокулярных клеток по каким-то причинам метаболически более активны, чем бинокулярные. Мы положили снимки в ящик стола и постарались забыть о них.
Прошло несколько лет, прежде чем нам или кому-либо другому довелось изучать с помощью той же реакции срезы первичной зрительной коры, параллельные ее поверхности. Когда это приблизительно одновременно было, наконец, сделано двумя группами (Анитой Хендриксон и А. Хамфри в Сиэтле и Дж. Хортоном и мной в Бостоне), выявился — ко всеобщему полному изумлению — узор, напоминающий материал в горошек. Пример показан на рис. 129. Вместо полосок мы увидели похожие на пузырьки образования, которые ни с чем из известного ранее не связывались. Неоднородности Уонг-Райли называли всеми мыслимыми именами: точки, пуфы, мушки, пятнышки. Мы назвали их «пузырьками» [blobs — это слово наглядно, узаконено (оно есть даже в оксфордском словаре английского языка) и как будто раздражает наших конкурентов].

Рис. 128. На этом поперечном срезе стриарной коры видны слои, окрашенные для выявления фермента цитохромоксидазы. Более темные области в слоях 2 и 3 (вертикальные полоски вверху) — «пузырьки».
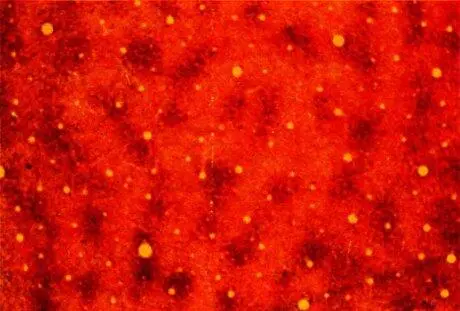
Рис. 129. Темные пятна представляют собой пузырьки, видимые «в плане»; около 50 таких пузырьков образуют характерный узор. Срез сделан через слой 3 параллельно поверхности коры на глубине около 0,5 мм. Срез проходит по границе между полями 17 (левая и средняя части) и 18 (правая часть), где пузырьков нет. (Желтые кружки — поперечные сечения кровеносных сосудов.)
Следующая задача была очевидной: мы должны были снова регистрировать ответы клеток в стриарной коре, гистологически контролируя эксперимент окраской на цитохромоксидазу, и попытаться выявить нечто особенное у клеток, находящихся в пузырьках. В 1981 году Маргарет Ливингстон и я принялись за эту работу. Результат оказался совершенно неожиданным. Проходя расстояние в четверть миллиметра, равное диаметру пузырька, можно исследовать примерно пять или шесть клеток. Всякий раз, когда мы пересекали пузырек, попадавшиеся на пути электрода клетки были полностью лишены ориентационной избирательности, что заметно контрастировало с высокой ориентационной избирательностью клеток, расположенных вне пузырьков.
Этому отсутствию ориентационной специфичности можно было бы дать два объяснения. Во-первых, эти клетки могли неизбирательно получать входные сигналы от соседних ориентационных клеток, лежащих вне пузырьков, и поэтому еще способны были отвечать специфической реакцией на линии (полоски и т.п.), но после объединения всех возможных ориентаций всякое предпочтение какой-либо из них исчезало. Во-вторых, они могли быть похожими на клетки коленчатого тела или на корковые клетки слоя 4C и, таким образом, быть проще внепузырьковых ориентационно-избирательных клеток. Вскоре вопрос был разрешен: оказалось, что большинство этих клеток имеет рецептивные поля с центром и периферией. Несколько дополнительных экспериментов убедили нас в том, что многие из них участвуют в кодировании цвета.
Больше половины пузырьковых клеток имели оппонентно-цветовые рецептивные поля с центром и периферией, но вели себя явно более сложным образом, чем клетки типа 1 наружного коленчатого тела. Они практически не отвечали на белые пятна любых размеров и формы. Но на небольшие цветные пятна, вспыхивающие в центре рецептивного поля, они энергично реагировали в одном диапазоне длин волны и затормаживались в другом диапазоне; некоторые активировались длинными волнами (красным светом) и подавлялись короткими (зеленым и синим светом), другие вели себя противоположным образом. Как и среди клеток коленчатого тела, мы могли, в зависимости от положения максимальных реакций на спектральной шкале, выделить два класса — красно-зеленые и сине-желтые клетки. (Здесь, как и ранее, слова «красный», «зеленый» и «синий» указывают на соответствующие типы колбочек, а слово «желтый» — на параллельные входы от красных и зеленых колбочек.) Таким образом, эти клетки весьма напоминали оппонентно-цветовые клетки коленчатого тела, обладающие только центром (тип 2). Но в отличие от клеток типа 2 эти цветокодирующие пузырьковые клетки чаще всего не отвечали на большие белые или цветные пятна, каков бы ни был спектральный состав света. Они вели себя так, как если бы центральная система рецептивного поля была у них окружена кольцом оппонентности. Если говорить о наиболее распространенном типе клеток, то центр типа r +g –, казалось, был окружен у них кольцом типа r –g +.
Мы с Маргарет Ливингстон назвали эти клетки двойными оппонентными из-за их красно-зеленой или желто-синей оппонентности в центре и антагонизма периферии по отношению к любой реакции в центре, будь она типа on или off. Поэтому они не отвечают не только на белый свет в любой геометрической конфигурации, но и на большие пятна, независимо от их спектрального состава. Центры рецептивных полей, как и у клеток типа 2, были в несколько раз больше, чем у клеток коленчатого тела типа 1. Как уже упоминалось, Н. Доу ввел термин двойные оппонентные для клеток, обнаруженных им в сетчатке золотой рыбки. Он предполагал, что сходные клетки могли бы участвовать в цвето-пространственных взаимодействиях у человека, и несколько лет спустя совместно с А. Пирлменом он старательно, хотя и безуспешно, искал такие клетки в наружном коленчатом теле макака.
В конце 60-х годов и позже двойные оппонентные клетки изредка находили в коре обезьян, но их не связывали четко с какими-либо анатомическими структурами. Нам до сих пор непонятны некоторые из их особенностей. Например, у только что описанных клеток типа r +g –красное пятно, окруженное зеленым, часто вызывает слабую реакцию вместо энергичной, которой можно было бы ожидать.
Читать дальшеИнтервал:
Закладка:







![Дэвид Доу - Почини свой мозг. Программа восстановления нейрофункций после инсульта и других серьезных заболеваний [litres]](/books/1143582/devid-dou-pochini-svoj-mozg-programma-vosstanovlen.webp)

