Дэвид Хьюбел - Глаз, мозг, зрение

- Название:Глаз, мозг, зрение
- Автор:
- Жанр:
- Издательство:Мир
- Год:1990
- Город:Москва
- ISBN:5-03-001254-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Дэвид Хьюбел - Глаз, мозг, зрение краткое содержание
В книге известного американского нейрофизиолога, лауреата Нобелевской премии, обобщены современные представления о том, как устроены нейронные структуры зрительной системы, включая кору головного мозга, и как они перерабатывают зрительную информацию. При высоком научном уровне изложения книга написана простым, ясным языком, прекрасно иллюстрирована. Она может служить учебным пособием по физиологии зрения и зрительного восприятия.
Для студентов биологических и медицинских вузов, нейрофизиологов, офтальмологов, психологов, специалистов по вычислительной технике и искусственному интеллекту.
Глаз, мозг, зрение - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
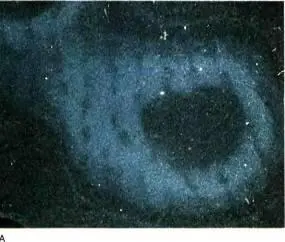
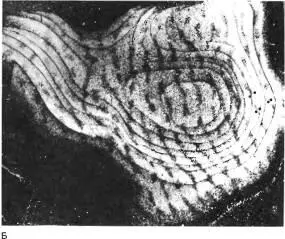
Рис. 145. Срезы зрительной коры макака, правый глаз которого был закрыт с момента рождения до 18-месячного возраста. В левый глаз инъецировали радиоактивную аминокислоту, и неделю спустя приготовили срезы мозга, параллельные поверхности зрительной коры. Кора имеет куполообразную форму, поэтому такие срезы вначале идут тангенциально, а затем образуют кольца (А) все большего диаметра, подобные кольцам луковицы. Эти кольца были вырезаны из микрофотографий и склеены вместе (Б). Впоследствии мы научились расправлять кору перед ее замораживанием, и тогда уже не приходилось разрезать и склеивать фотографии серийных срезов. На обычном радиоавтографе зерна серебра выглядят черными на белом фоне. Мы, однако, применяли метод исследования в темном поле, при котором зерна серебра рассеивают свет и их скопления выглядят как светлые участки. Светлые полоски — места поступления в слой 4C меченых атомов из открытого глаза — расширены, а темные (связанные с закрытым глазом) сильно сужены.

Рис. 146. Другой обезьяне метку ввели в закрытый глаз. Срез сделан в поперечном, а не в тангенциальном направлении. На этом снимке, полученном в темном поле, светлые полоски в слое 4C (которые видны здесь в поперечном сечении) сильно сужены.
Результаты, полученные на слое 4C, подкрепили наши сомнения относительно конкурентной модели, связанные с фактом атрофии клеток коленчатого тела: либо гипотеза о конкуренции была неверна, либо какая-то погрешность была в наших рассуждениях. Оказалось, что погрешность касалась как коленчатого тела, так и коры. В коре нашей ошибкой было предположение, что к моменту, когда мы закрывали глаза новорожденным животным, колонки глазодоминантности были уже вполне сформированы.
Очевидный способ получения сведений о колонках глазодоминантности у новорожденных животных заключался в проверке распределения волокон, входящих в слой 4C, путем инъекции метки в глаз на первый или второй день жизни. Результат оказался неожиданным. Вместо четких, ясно очерченных полосок в слое 4C мы наблюдали непрерывное распределение метки. Радиоавтограф слева на рис. 147 показывает этот слой в поперечном разрезе, и мы не видим никаких признаков колонок. Только если мы нареза́ли кору параллельно ее поверхности, можно было увидеть слабую волнистость с полумиллиметровыми интервалами, как это показано на радиоавтографе справа. Очевидно, волокна, растущие от коленчатого тела в кору, не разветвляются сразу же к областям, связанным с левым и с правым глазом. Они сначала посылают свои ветви повсюду в радиусе нескольких миллиметров и лишь позднее, приблизительно к моменту рождения животного, втягиваются обратно и окончательно распределяются. Слабая волнистость у новорожденного показывает, что втягивание началось еще до рождения; Пашко Ракич с большими трудностями сумел инъецировать метку в глаз утробного плода обезьяны и выяснил, что оно начинается за несколько недель до рождения. Инъецируя метку в один глаз в различные моменты после рождения, мы смогли легко установить, что в первые две или три недели в слое 4 происходит постепенное втягивание окончаний, так что к четвертой неделе формирование полосок завершается. Узор полосок и их периодичность с шагом 0,8 миллиметра, таким образом, являются врожденными.
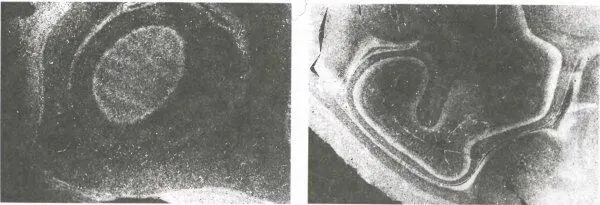
Рис. 147. Слева: поперечный срез слоя 4C коры новорожденного макака после инъекции радиоактивной метки в глаз. Снимок сделан в темном поле, поэтому меченые участки выглядят светлыми. Их непрерывность показывает, что входы от каждого глаза не сгруппированы в полоски, а перемешаны во всем слое. (Белая полоса между слоями 4C, находящимися у поверхности мозга и в глубине, представляет собой белое вещество мозга, заполненное волокнами с меткой, идущими от наружных коленчатых тел.)
Справа: здесь показан срез другого полушария; при изготовлении среза нож слегка коснулся участка стриарной коры, находящегося в глубине. В верхней части слоя 4C можно усмотреть намек на полоски. (Эти полоски находятся в подразделении, связанном с крупноклеточными слоями коленчатого тела.) Более глубинный подслой β образует непрерывное кольцо вокруг α и поэтому предположительно позднее дифференцируется по глазодоминантности.
Мы легко подтвердили представление о постнатальном втягивании окончаний, регистрируя ответы слоя 4C обезьян вскоре после рождения. По мере продвижения электрода вдоль этого слоя параллельно его поверхности мы везде смогли вызвать активность от обоих глаз вместо четкого чередования глаз, наблюдаемого у взрослых животных. Карла Шатц продемонстрировала аналогичный процесс развития коленчатого тела у кошки: у утробного плода многие клетки коленчатого тела временно получают входы от обоих глаз, но утрачивают один из входов по мере образования слоистой структуры. Таким образом, и в коре, и в коленчатом теле мы имеем примеры формирования и последующей спонтанной дегенерации синапсов в процессе развития.
Окончательная картина чередования полос для левого и правого глаза в корковом слое 4C развивается нормально даже в том случае, если оба глаза закрыты путем сшивания век; это означает, что надлежащие связи могут создаваться и без участия индивидуального опыта. Мы полагаем, что в процессе развития входные волокна от обоих глаз конкурируют в слое 4C таким образом, что если входы от одного из глаз в каком-то месте преобладают, то преимущество этого глаза имеет тенденцию увеличиваться, а число входов от другого глаза соответственно убывает. Даже небольшое начальное неравенство при этом стремится постепенно возрастать до тех пор, пока всюду в слое 4 в возрасте одного месяца не появятся отчетливые полоски с полным доминированием. При закрытом глазе баланс нарушается, и на границах полосок, где в норме исход борьбы решает прямое столкновение, открытый глаз получает преимущество и одерживает верх, что показано на рис. 148.
Мы не знаем, что́ при нормальном развитии приводит к начальному дисбалансу, но при столь неустойчивом равновесии дело может решить даже мельчайшее различие. Почему развивающийся узор должен состоять из параллельных полосок шириной 0,5 мм каждая, остается предметом догадок. Некоторые исследователи предполагают, что аксоны от одного и того же глаза, сближаясь, начинают притягивать друг друга, а от разных глаз — отталкивать друг друга, причем на малых расстояниях отталкивание слабее притяжения, так что притяжение побеждает. С увеличением расстояния притягивающая сила убывает быстрее отталкивающей, так что на некотором удалении побеждает отталкивание. Шириной зоны этих конкурентных взаимодействий и определяются размеры колонок. Согласно математическим расчетам, для получения параллельных полосок (в противовес шахматному порядку или островкам аксонов от левого глаза в сплошной массе аксонов от правого глаза) требуется лишь то, чтобы границы между колонками были как можно короче. Итак, мы имеем возможность объяснить сужение и расширение колонок, показав, что в то время, когда глаз был закрыт в раннем периоде жизни, конкуренция все же была возможной.
Читать дальшеИнтервал:
Закладка:







![Дэвид Доу - Почини свой мозг. Программа восстановления нейрофункций после инсульта и других серьезных заболеваний [litres]](/books/1143582/devid-dou-pochini-svoj-mozg-programma-vosstanovlen.webp)

