Станислав Дробышевский - Шанс для приматов

- Название:Шанс для приматов
- Автор:
- Жанр:
- Издательство:сет. публикация
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Станислав Дробышевский - Шанс для приматов краткое содержание
Главы из книги известного российского антрополога, посвященные древним млекопитающим. Описываются разные их группы и оценивается потенциальная их возможность пойти по пути увеличения и усложнения мозга. Что позволило нашим далеким предкам использовать свой шанс и превратиться в нас с Вами, и что помешало другим группам млекопитающих сделать это…
Шанс для приматов - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Пикродонтиды Picrodontidae, с их морщинистыми многобугорковыми, очень широкими — особенно первыми — и совсем не стёртыми молярами, стали очень специализированными фруктоядами, а может, даже питались преимущественно нектаром, пыльцой и древесным соком. Череп Zanycteris резко сужался спереди, этим пикродонтиды были похожи на современных мышиных лемуров Cheirogaleinae и хоботноголовых кускусов Tarsipedidae, питающихся нектаром. Пикродонтиды, видимо, дальше всех из плезиадапиформов ушли от исходной насекомоядности. Забавно, что зубы пикродонтид столь необычайно похожи на зубы летучих мышей, что в 1935 году Дж. Симпсон даже предполагал, что пикродонтиды относятся к рукокрылым (Simpson, 1935).
Паромомиоиды Paromomyoidea имели квадратные уплощённые моляры, приспособленные для перетирания фруктов, впрочем, в диете немалую роль могли играть древесный сок и насекомые. В последовательном ряду палеоценовых родов наблюдается быстрое удлинение и истончение нижних резцов с появлением промежутка — диастемы — между резцами и премолярами. На основании строения изолированных костей конечностей предполагалось, что паромомииды имели летательную перепонку, как шерстокрылы, но обнаружение и изучение целых скелетов показало, что эти звери были больше похожи на белок, а не на летяг.
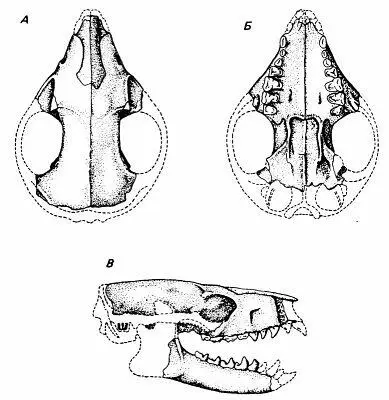
Череп паромомиоидового примата Palaechthon (средний палеоцен)
Все эти специализированные плезиадапиформы, конечно, не были предками приматов, но известны и более генерализованные их варианты, например, средне-верхнепалеоценовые североамериканские палеохтониды Palaechthonidae. У них сохранялась исходная для приматов зубная формула, не было диастемы между резцами и премолярами, отсутствовали гипертрофии каких-либо зубов. Строение черепа Palaechton nacimienti было самым примитивным среди всех плезиадапиформов: глазницы были маленькие, ориентированы вбок и широко расставлены, заглазничное сужение не выражено, обонятельные луковицы большие; судя по увеличенному подглазничному отверстию, имелись длинные вибриссы. Ориентировались палеохтониды в основном с помощью осязания, слуха и обоняния, а не зрения. Судя по всему, вели они в основном наземный образ жизни. В немалой степени все эти черты были связаны с малыми размерами тела и насекомоядностью палеохтонид (Kay et Cartmill, 1977). Показательно, что группа палеохтонид — вовсе не древнейшая среди плезиадапиформов; с одной стороны, она сохраняла исходную насекомоядность и экологически схожа с тупайями, с другой — по времени близка к настоящим приматам границы палеоцена и эоцена. Вряд ли настоящих приматов дали палеохтониды с их особенностями, но это должны были быть очень похожие на палеохтонид существа, только более древесные и всеядные.
Насекомоядность вполне сочеталась с древесностью у микромомиид Micromomyidae, которые были самыми мелкими плезиадапиформами — намного меньше современных мышиных лемуров. При их размере другая диета, нежели насекомые, млекопитающим резко противопоказана. Эти крошечные зверюшки жили в конце палеоцена и начале эоцена в Северной Америке.
Позднейшей группой насекомоядных плезиадапиформов были толиапиниды Toliapinidae (Hooker et al., 1999). Они появились лишь в конце палеоцена и дожили до среднего эоцена. Большинство видов, включая примитивнейшего Sarnacius gingerichi, найдено в Европе, но уже в начале палеоцена Altiatlasius koulchii мигрировал в Марокко, а самый поздний — Seia shahi — обнаружен в Пакистане. Altiatlasius koulchii первоначально был описан как древнейший настоящий примат семейства Omomyidae, но позже пересмотрен как плезиадапиформ. Толиапиниды были весьма схожи с микромомиидами по образу жизни и являются их полным "старосветским" аналогом; к сожалению, от тех и других найдено очень немного останков.
Плезиадапиформы имели мозг гораздо меньший, чем у приматов таких же размеров, но бóльший, чем у других зверей. Возможно, важнейшим признаком, обеспечившим в далёком будущем и наше появление, стало соотношение жевательной мускулатуры и мозговой коробки. Жевательные мышцы крепятся у млекопитающих на сагиттальном гребне, тянущемся по верху мозговой коробки; таким образом, они окружают её, уже с младых ногтей своим давлением способствуют раннему зарастанию швов черепа и препятствуют росту мозга. У приматомофов, начиная с плезиадапиформов, стала усиливаться тенденция к ослаблению обоняния и развитию стереоскопичности зрения. Это привело к повороту глазниц вперёд, уменьшению обонятельных луковиц и решётчатой кости, а в итоге — появлению заглазничного сужения черепа в том месте, где у всех приличных зверей находится обонятельный мозг (для гоминид сильное сужение — примитивный признак, а для приматов как отряда — прогрессивный, таковы причуды эволюции).
Таким образом, значительная часть сагиттального гребня оказалась впереди от мозговой коробки, а коробка, соответственно, сдвинулась от мышц назад, освободившись от его гнёта и получив возможность увеличиваться более долгий срок в онтогенезе. Фактически, редукция мозга привела к росту мозга!
Парадокс, но факт.
Впрочем, прогрессивные особенности плезиадапиформов не уберегли их от вымирания. Традиционно и по-умолчанию считалось, что в их исчезновении повинны более продвинутые настоящие приматы. Однако изучение частот встречаемости останков рисует другую картину (Fleagle, 1999). Резкий спад численности плезиадапиформов начинается совершенно синхронно со столь же резким подъёмом численности первых грызунов, тогда как лишь через некоторое время появляются и начинают распространяться полуобезьяны. Стало быть, плезиадапиформы проиграли эволюционную гонку грызунам, зато своим исчезновением освободили дорогу полуобезьянам. Таким образом, отличия плезиадапиформов от приматов оказываются крайне существенными: то, чего не было у первых, не позволило им успешно конкурировать с грызунами; то же, что приобрели вторые, вывело их на новый уровень, недоступный грызунам.
Среди плезиадапиформов особняком стоит семейство Adapisoriculidae. Особенности его представителей позволяли разным исследователям включать его в сумчатых, насекомоядных, лептиктид или миксодектид, а также считать предковым для тупайй. Адаписорикулиды назывались и как возможные предки афросорицид Afrosoricida, включающих современных златокротов Chrysochloridae и тенреков Tenrecidae (Seiffert, 2010). Несмотря на существенные различия с плезиадаписовыми в зубной системе, строение посткраниального скелета у плезиадапиформов, шерстокрыловых и адаписорикулид весьма схоже (Smith et al., 2010); судя по всему, это были древесные насекомоядные животные. Более того, плечевая кость адаписорикулид имеет промежуточное строение между вариантами меловых "кондиляртр" и кайнозойских эуархонт, причём в ряду самих адаписорикулид обнаруживается постепенный переход от более примитивного варианта мелового Deccanolestes к более эуархонтовому у палеогеновых форм (Boyer et al., 2010b). Если считать адаписорикулид приматами или приматоморфами, а Deccanolestes hislopi считать адаписорикулидом, то эта группа становится самой древней среди приматоморфов, поскольку указанный вид обнаружен в позднемеловых отложениях Индии.
Читать дальшеИнтервал:
Закладка:

![Станислав Дробышевский - Палеонтология антрополога. Книга 1. Докембрий и палеозой [litres]](/books/1073953/stanislav-drobyshevskij-paleontologiya-antropologa.webp)
