Александр Уголев - Естественные технологии биологических систем

- Название:Естественные технологии биологических систем
- Автор:
- Жанр:
- Издательство:Наука
- Год:1987
- Город:Ленинград
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Уголев - Естественные технологии биологических систем краткое содержание
Книга посвящена концепции естественных технологий живых систем на различных уровнях организации последних и изложению доказательств, позволяющих преодолеть противопоставление естествознания и технологии. Эта концепция обосновывается на примере наиболее важных процессов в живых системах, их эволюции и происхождения. Охарактеризованы некоторые закономерности, которые могут быть интерпретированы как общие для естественных технологий живой природы и производственных технологий. Показано, что такие подходы плодотворны для понимания биологии в целом, процессов, протекающих в живых системах различной сложности, взаимодействий естественных и производственных технологий, в частности в медицине, экологии, питании и т.д.
Естественные технологии биологических систем - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Попытаемся показать, что многообразные высокоразвитые формы управления в сложных организмах достигаются благодаря комбинации стандартных функциональных блоков, образующих специализированные и приспособленные для определенных задач цепи.
Химические сигналы.Первоначально предполагалось, что каждый эффект и каждый источник физиологически активного вещества связан с особым гормоном. При действии на мишень нескольких гормонов принималось, что лишь один из них оказывает физиологическое действие, а другие — фармакологическое. Однако множественный контроль клеток различных типов пищеварительной системы известен. Например, обмен воды между тканями тонкой кишки и ее полостью контролируется многими гормонами (гастрин, секретин, холецистокинин, ВИП, ГИП, вазопрессин, субстанция Р, бомбезин, серотонин и др.), продуцируемыми эндокринными клетками как желудочно-кишечного тракта, так и другими эндокринными органами, а также простагландинами. Чувствительность к различным гормонам и другим физиологически активным веществам означает наличие рецепторов многих типов или сложных рецепторов, способных связывать не один, а два или более видов лигандов. (Под рецепторами понимается большая группа структур, обладающих высоким сродством и способностью связываться с определенными типами лигандов). Можно ли считать, что рецепторные блоки различных органов идентичны или они специфичны для каждого соответствующего органа? Мембранные рецепторы являются гликопротеинами, и варьирование их свойств может зависеть от уровня посттрансляционного гликозилирования. В то же время существуют данные в пользу сходства или идентичности одноименных рецепторов в различных органах. В частности, это показано для таких ранее казавшихся локальными факторов, как субстанция Р, холецистокинин, ВИП, гастрин и др.
Внутриклеточные сигналы.На уровне сигнальных рецепторных блоков концепция их универсальности и специализации на основе рекомбинации находит новое подтверждение. Например, секреторные процессы в ацинарных клетках поджелудочной железы контролируются системой нервных и гормональных сигналов. В частности, выброс секреторных гранул происходит под влиянием холецистокинина и ацетилхолина. Холецистокинин длительное время рассматривался как специфический стимулятор секреции ферментов. Однако позднее было обнаружено, что этот механизм не специфичен, а включен в систему вторичных мессенжеров — цАМФ и Са 2+. Так, холецистокинин контролирует многие цАМФ- и Са 2+-зависимые процессы, в том числе внутренней секреции, стимулируя выделение ряда гормонов, и действует аналогично медиатору. Многие гормоны выполняют также функции нейротрансмиттеров. Эту роль могут играть гастрин и холе-цистокинин; серотонинподобные вещества, вызывающие возбуждение в межнейронных синапсах; АТФ — скорее всего в тормозящих нейронах; субстанция Р, выполняющая функции возбуждающего, а энкефалин и соматостатин — тормозящего нейротрансмиттеров. Предполагается также нейротрансмиттерная функция ВИПа (рис. 29).
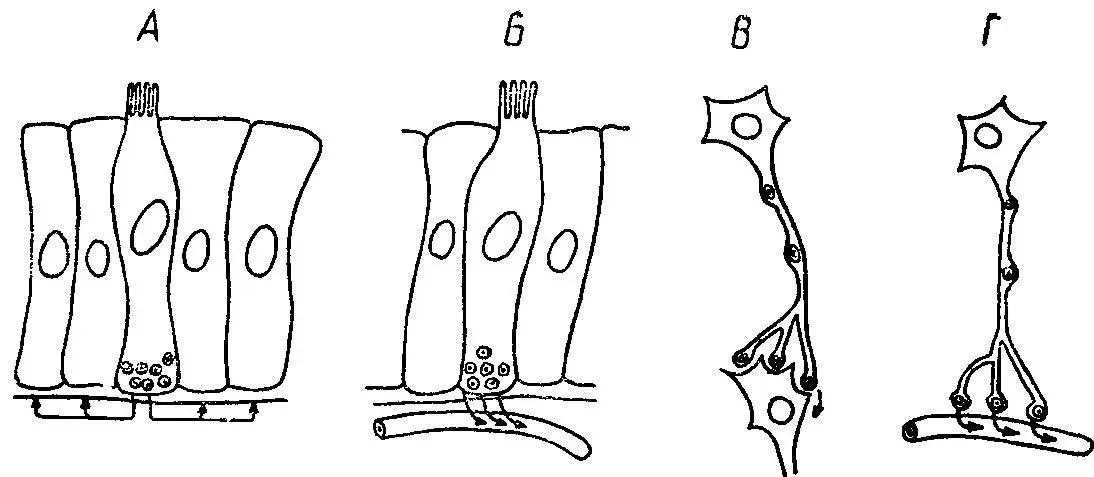
Рис. 29. Схема способа передачи пептидов, действующих в качестве паракринных мессенжеров (А), кишечных гормонов ( Б ), нейротрансмиттеров (В) и нейрогормонов (Г).
Те же самые молекулы могли бы функционировать в каждой из систем одного и того же организма.
Принцип работы рецепторных блоков можно проиллюстрировать на примере рецепторно-аденилатциклазного комплекса. Такой комплекс осуществляет передачу сигналов путем их ретрансляции с помощью аденилатциклазы. Сущность процесса сводится к образованию комплекса рецептор—гормон, что приводит к стимуляции активности аденилатциклазы, локализованной на внутренней стороне мембраны. Активация аденилатциклазы первичным мессенжером связана с его взаимодействием с рецептором, каталитической и, возможно, промежуточной субъединицами фермента. При стимуляции активности фермента происходит увеличение образования цАМФ, что вызывает цепную реакцию с отрицательной обратной связью, приводящую к ее выключению. Ниже приведен список гормонов, стимулирующих или подавляющих активность аденилатциклазы (табл. 11).
Таблица 11
Гормоны, оказывающие влияние на аденилатциклазную активность различных тканей
| Гормон | Ткань-мишень |
| Катехоламины | Многие ткани |
| Глюкагон | Печень, жировая ткань, В-клетки поджелудочной железы |
| АКТГ (адренокортикотропный гормон) | Кора надпочечников, жировая ткань |
| ТСГ (гормон, стимулирующий выделение тиреотропина) | Щитовидная железа |
| Паратиреоидный гормон | Почки, костная ткань |
Ясно, что с помощью различных соотношений стандартных рецепторных и циклазных блоков может быть достигнута высокая избирательность регуляции различных органов (рис. 30). Вместе с тем при некоторых условиях даже небольшое количество определенных рецепторов может быть важным при развитии адаптационных (например, адаптационно-компенсаторных) реакций или при развитии патологических процессов, например вследствие побочных эффектов гормональных веществ.
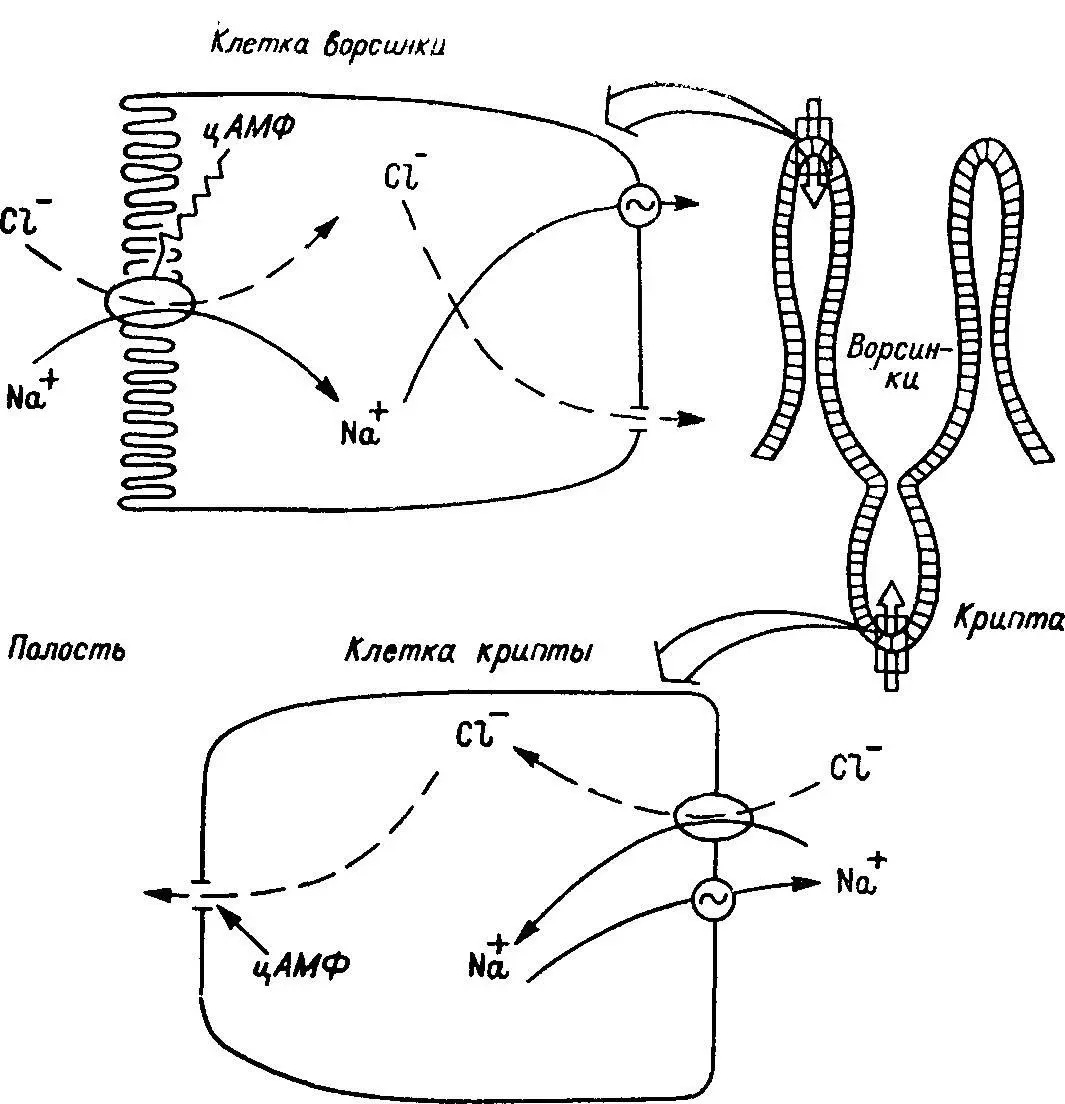
Рис. 30. Предполагаемая схема действия цАМФ на транспорт ионов в ворсинках и криптах тонкой кишки млекопитающих.
Показаны два секреторно-чувстительных процесса, разделенных пространственно: абсорбтивный электронезависимый, локализованный в клетках ворсинок, и электрогенный секреторный, локализованный в клетках крипт. Эта же схема иллюстрирует предполагаемую цАМФ-медиироваввую секрецию электролитов в тощей кишке. Антиабсорбтивные (в клетках ворсинок) и секреторные (в клетках крипт) эффекты ввутриклеточных мессенжеров разделены. Гипотеза может иметь отношение ко всей секреции, стимулируемой циклическими нуклеотидами и Са 2+, так как цАМФ и Са 2+-ионофоры оказывают эффект на транспорт, сходный с эффектами цАМФ.
Таким образом, наряду с воспринимающим блоком — собственно рецептором — существует эффекторный блок — аденилатциклаза. Эта система универсальна и присутствует в клетках всех органов и тканей. Система циклических нуклеотидов имеется не только у позвоночных, большинства беспозвоночных, включая первичноротых, но и у одноклеточных организмов. У последних, так же как и у грибов, цАМФ выполняет роль передатчика сигналов не внутрь клеток, а от одной особи к другой. Следовательно, цАМФ первично возникает на весьма ранних стадиях формирования эукариотов как типичный гормон и, сохраняясь как химический мессенжер и постепенно подвергаясь интернализации, превращается во вторичный мессенжер.
Читать дальшеИнтервал:
Закладка:









