Александр Уголев - Естественные технологии биологических систем

- Название:Естественные технологии биологических систем
- Автор:
- Жанр:
- Издательство:Наука
- Год:1987
- Город:Ленинград
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Уголев - Естественные технологии биологических систем краткое содержание
Книга посвящена концепции естественных технологий живых систем на различных уровнях организации последних и изложению доказательств, позволяющих преодолеть противопоставление естествознания и технологии. Эта концепция обосновывается на примере наиболее важных процессов в живых системах, их эволюции и происхождения. Охарактеризованы некоторые закономерности, которые могут быть интерпретированы как общие для естественных технологий живой природы и производственных технологий. Показано, что такие подходы плодотворны для понимания биологии в целом, процессов, протекающих в живых системах различной сложности, взаимодействий естественных и производственных технологий, в частности в медицине, экологии, питании и т.д.
Естественные технологии биологических систем - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Ряд клеток характеризуется тем, что универсальные транспортные блоки расположены на клеточной поверхности иначе. В частности, в ацинарных клетках поджелудочной железы, осуществляющих внешнюю секрецию ферментов, насосы и Na +-зависимые транспортеры расположены в базолатеральной мембране. Этого достаточно для обеспечения градиента Na +между вне- и внутриклеточной жидкостями и поступлением нутриентов из крови в цитоплазму, а также для энергизации многих других процессов. Кроме того, локализация насосов и каналов в базолатеральной мембране исключает интенсивное поступление потоков воды и солей в апикально-базальном направлении.
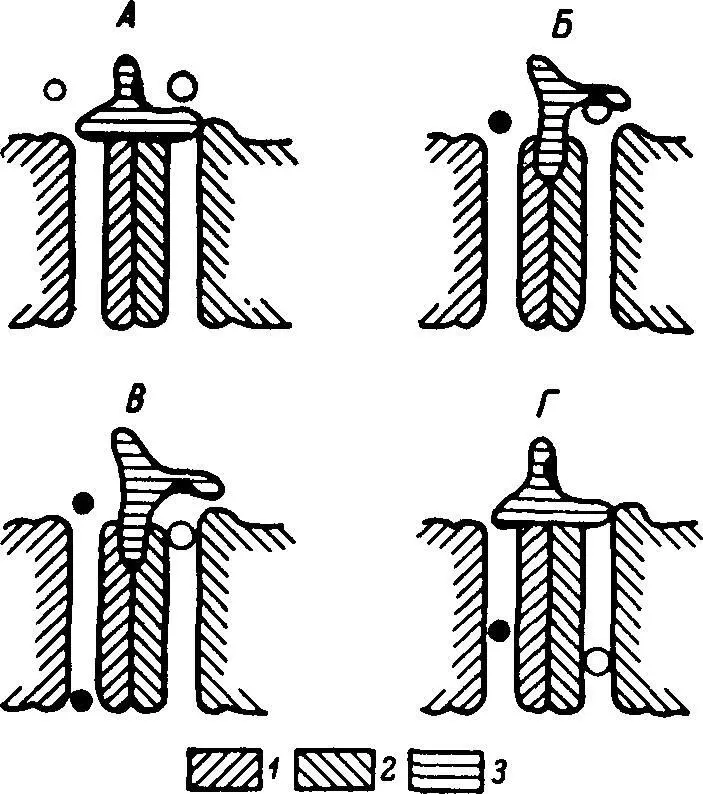
Рис. 28. Схема действия транспортера глюкозы и натрия в плазматической мембране.
А — исходное состояние транспортера: оба канала закрыты; Б — после связывания глюкозы с аллостерическим центром на воротном устройстве: натриевый канал открылся; В — при прохождении натрия по натриевому каналу произошла аллостерическая активация глюкозного канала и молекула глюкозы перешла с воротного устройства на вход своего канала; Г — освобождение центра на воротном устройстве влечет за собой закрытие натриевого и глюкозного каналов. 1 — натриевый канал; 2 — глюкозный канал; 3 — воротное устройство. Светлые пружки — молекулы глюкозы, черные — ионы натрия.
Согласно современным представлениям, при Na +-зависимом транспорте глюкозы через плазматическую мембрану клеток эукариотов происходит образование тройного комплекса: Na +—глюкоза—транспортер. Однако более адекватной представляется наша модель транспортера (рис. 28). Она характеризуется наличием двух параллельно взаимодействующих каналов (для Na +и для глюкозы) и поверхностного (воротного) белка, связывающего глюкозу на входе в транспортную систему. Транспортный цикл такой модели состоит в следующем: 1) исходно глюкозный и натриевый каналы неактивны; 2) при связывании глюкозы с аллостерическим центром на воротном белке натриевый канал активируется (эта стадия напоминает контроль натриевой проницаемости ацетилхолином) и Na +движется из экстра- в интрацеллюлярную жидкость; 3) на определенном этапе движения Na +по каналу происходит аллостерическая активация глюкозного канала; 4) через активированный канал транспортируется молекула глюкозы, первоначально фиксированная в воротном устройстве; 5) освобождение воротного устройства сопровождается дезактивацией натриевого канала. Реактивация последнего происходит при связывании с аллостерическим центром следующей молекулы глюкозы. При температуре 16° С глюкозный канал инактивируется, а натриевый продолжает функционировать. Из нашей модели вытекает, что скорость транспорта глюкозы и Na +в определенном диапазоне зависит от концентрации Na +во внеклеточной жидкости. Важно, что Ка +-зависимые транспортеры глюкозы и аминокислот, поскольку показана их идентичность в клетках различных типов и у организмов разных групп, должны соответствовать одной из моделей молекулярных машин.
Насколько можно судить, все разнообразие процессов, связанных с мембранной проницаемостью электролитов и их мембранным транспортом (по крайней мере у всех эукариотов), осуществляется с помощью тех же стандартных функциональных блоков, которые описаны выше. Поразительно, но «примитивные» функции низших организмов и совершенные функции высших организмов реализуются близкими или идентичными функциональными блоками. На уровне элементарных функций организмы, занимающие крайнее положение на вершине и у основания эволюционной лестницы, трудно различимы. Одноименные транспортеры органических субстанций у животных разных видов, по-видимому, также близки или сходны по своей молекулярной структуре и принципам функционирования, хотя этот вопрос изучен меньше, чем в отношении АТФаз. В пользу большой близости этого типа функциональных блоков говорит сходство, а иногда идентичность реакций различных транспортеров на действие ингибиторов и активаторов.
Несмотря на то что сравнительная физиология ионных каналов изучена недостаточно, существуют данные о значительном сходстве их молекулярной структуры и функциональной организации. Вероятно, у млекопитающих и насекомых имеются все основные типы ионных каналов — натриевые, калиевые, хлорные и др.
Далее, у млекопитающих, рыб и насекомых продемонстрировано наличие специализированных натриевых каналов, чувствительных к амилориду, а также каналов, реализующих котранспорт Na +и Сl -, чувствительных к фуросемиду. Интересно, что те же самые ингибиторы специфически тормозят транспорт ионов в мальпигиевых канальцах и толстой кишке насекомых. Недавно выявлен котранспортер, осуществляющий котранспорт в клетку одного иона Na +, одного иона К +и двух ионов Сl -. Этот механизм обнаружен в клетках почки и тонкой кишки млекопитающих, в клетках различных органов рыб и насекомых.
До недавнего времени считалось, что функции секреции, экскреции и всасывания имеют глубокие различия, касающиеся механизмов функционирования и эволюционного происхождения. Однако между этими процессами принципиальной разницы не существует. Все основные функциональные блоки, упомянутые выше, лежат и в основе перечисленных функций, в ряде случаев полярных по своему физиологическому значению (например, всасывание и экскреция).
Характеристики ферментных и транспортных систем клеток, выполняющих различные функции (например, всасывание в тонкой кишке, реабсорбция воды в протоках пищеварительных желез и т.д.), очень близки. В апикальной мембране клеток тонкой кишки и почечных канальцев присутствуют трансмембранные ферменты, Ка +-зависимые транспортеры глюкозы, аминокислот и других нутриентов. В базолатеральной мембране этих клеток локализованы близкие или идентичные по молекулярной структуре АТФазы. Системы с множеством транспортеров, локализованных в апикальной мембране, и натриевых насосов, расположенных в базолатеральной мембране, распространены очень широко. В некоторых структурах они выполняют не только резорбтивную, но и секреторную функцию.
Существенно, что при электронной микроскопии многие субклеточные и молекулярные структуры клеток тонкой кишки человека и высших животных неотличимы от таковых клеток кишки рыб, насекомых, моллюсков, круглых червей и т.д., а клетки тонкой кишки — от клеток плаценты, канальцев почек и других клеток.
5.5. Регуляторные блоки
В течение долгого времени постулировалась специфичность регуляторных сигналов и рецепторов, воспринимающих эти сигналы. В отношении химических сигналов идея специфичности достигла наибольшего развития. Действительно, эти сигналы, действующие через кровь, должны находить свои клетки-мишени и оказывать на них определенный эффект. В то же время число сигнальных молекул по сравнению с тем количеством, которое следовало бы ожидать, исходя из специфичности химических сигналов, невелико. Рецепторы для каждого типа сигнальных молекул локализованы практически в клетках всех типов. Кроме того, на примере кишечной гормональной системы продемонстрировано, что клетки, продуцирующие казавшийся ранее уникальным гормон, могут быть дублированы или даже мультиплицированы.
Читать дальшеИнтервал:
Закладка:









