Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
В таких условиях появление основанного на РНК механизма трансляции привело бы к производству репликазы и, с последующей репликацией РНК, к критическому переходу от антропной причинности к биологическому отбору (см. рис. 12-6). В принципе начало биологической эволюции можно себе представить и с репликазой как единственным изначально активным белком. Однако, учитывая вероятное существование реакторов, производящих РНК, которые обсуждались ранее в этой главе, вполне возможно, что с появлением трансляции другие случайные последовательности РНК послужили основой предковых форм наиболее распространенных белковых укладок, порождая несколько белковых функций (например, РНК-связывающие белки и примитивные ферменты, катализирующие синтез нуклеотидов) и тем самым придавая минимально необходимую устойчивость формирующейся биологической системе. Появление этих укладок можно назвать Большим взрывом белковой эволюции.
Как уже отмечалось, современный универсальный генетический код гораздо более надежен, чем был бы случайный, по отношению к мутационным и, вероятно также, к трансляционным ошибкам. Эта устойчивость проявляется и в очевидной неслучайности структуры кода, выражающейся в первую очередь в том, что серия кодонов, которые отличаются только третьей позицией, кодирует либо одну и ту же, либо две подобные аминокислоты, и в других особенностях соответствия кодонов аминокислотам (Koonin and Novozhilov, 2009). Примечательно, что предполагаемый предковый «дублетный» код, в котором третья позиция не несла никакой информации, мог быть даже более надежным, чем современный (Novozhilov and Koonin, 2009). Надежность, как обычно предполагается, эволюционировала в ходе оптимизации кода. Однако модель АХЭ предлагает альтернативную точку зрения, при которой базовая структура кода возникла по чистой случайности, поскольку только коды с определенным минимальным уровнем надежности позволили бы функциональной репликазе появиться в прорывной системе. Конечно, этот сценарий не исключает и последующей корректировки кода биологической эволюцией, что, по всей вероятности, в действительности произошло.
Таким образом, гипотеза АХЭ снимает парадоксы происхождения репликации и трансляции предположением о том, что оба этих процесса, в их примитивных формах, не произошли в результате биологической эволюции, а возникли случайно, как связанная система, в соответствии с антропным принципом.
Гипотеза АХЭ, несомненно, должна показаться большинству эволюционных биологов нелепой и возмутительной, поскольку она уклоняется от поисков «механизмов» доклеточной эволюции. Тем не менее существуют смягчающие обстоятельства. Во-первых, постулируемая возможность возникновения сопряженной репликационно-трансляционной системы не требует никаких неизвестных процессов. Напротив, для нее необходимы только хорошо известные, обычные реакции, такие как полимеризация нуклеотидов и аминокислот и фосфорилирование/дефосфорилирование нуклеотидов; при этом нужны только распространенные в химии и биохимии взаимодействия. Как уже отмечалось в этой главе, элементарные реакции, необходимые для трансляции (активация аминокислот, аминоацилирование РНК и транспептидация), легко моделируются с помощью рибозимов, в противоположность репликации РНК, которой, как известно, трудно достичь в отсутствие белков. Во-вторых, исключая разве что полную неадекватность нынешнего понимания условий на первичной Земле, любые мыслимые сценарии эволюции жизни обязательно требуют сочетания маловероятных условий и событий до начала биологической эволюции. Список таких событий включает в себя абиогенный синтез довольно сложных и не очень устойчивых органических молекул, таких как нуклеотиды, накопление этих молекул в соответствующих ячейках до высоких концентраций и их полимеризацию с получением полинуклеотидов достаточного размера и разнообразия. Таким образом, независимо от космологических соображений, некоторые формы антропной причинности представляются неизбежным аспектом эволюции жизни (см. рис. 12-6).
Я привел сценарий АХЭ, чтобы показать, что диапазон сложности, открытой для антропной причинности, может быть гораздо шире, чем предполагалось ранее, настолько, что первичная репликационно-трансляционная система могла возникнуть без биологического отбора. Случайное происхождение изощренной системы, способной исполнять сложные биологические функции, может показаться бессмыслицей. Я полагаю, однако, что это всего лишь семантическая ловушка. До наступления биологической эволюции не может быть «функции», только сложность, а модель МММ гарантирует возникновение любого уровня сложности (это гарантированно произойдет «где-то» в бесконечной Вселенной, но антропный принцип прямо помещает эти события на Землю).
Все эти соображения провоцируют довольно-таки кошмарный вопрос: имеет ли какое-либо значение в бесконечно избыточном мире МММ биологическая эволюция вообще и дарвиновский отбор в частности? Разве не возникнет система любой, даже наивысшей, сложности просто случайно? Ответ: да, но этот вопрос упускает важное обстоятельство. В модели МММ случайное появление бесконечного числа сложных биот неизбежно, но несравнимо реже, чем развитие по сценарию АХЭ, которое включает в себя переход от случайности к биологической эволюции после стадии прорывной системы (см. рис. 12-6). Начало биологической эволюции канализирует исторический процесс, сокращая многочисленные траектории, которые возможны в принципе, до относительно малого числа стабильных и более вероятных – тех, что совместимы с дарвиновским режимом эволюции сложных систем (см. рис. 12-7). Этот переход приводит к гораздо большей скорости эволюционных изменений, чем та, что достигается случаем, и, как только возникает возможность для биологической эволюции, антропная причинно-следственная связь отходит на второй план в истории жизни. Конечно, «второй план» не означает потери важности: непредвиденные случайности имеют решающее значение, особенно на переходных этапах эволюции (см. обсуждение ранее в этой книге, особенно в гл. 7). Таким образом, в любой реконструкции происхождения жизни и ранней эволюции порог должен быть связан с самой нижней точкой, то есть минимально сложной системой, способной к биологической эволюции.
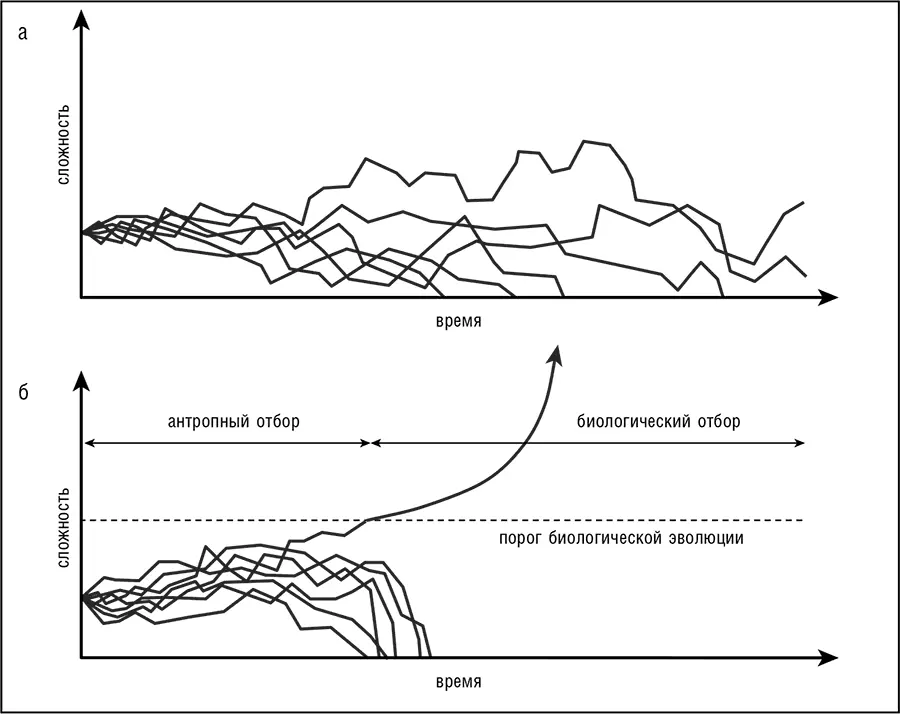
Рис. 12-7. Отсечение эволюционных траекторий на пороге биологической эволюции: а – только химическая эволюция; б – возникновение биологической эволюции.
Сильная форма гипотезы АХЭ, согласно которой прорывным этапом в истории жизни была примитивная совместная репликационно-трансляционная система (см. рис. 12-6), в принципе легко опровержима. Такая система должна рассматриваться как верхняя граница сложности для прорывного этапа. Как только возможность биологической эволюции при более низком уровне сложности, таком как мир РНК, будет убедительно продемонстрирована, а путь от мира РНК к системе трансляции очерчен либо опытным путем, либо в убедительной модели, сильная форма гипотезы АХЭ будет опровергнута. Демонстрация самостоятельного возникновения жизни на нескольких планетах в нашей Вселенной будет иметь тот же результат. В приложении приведен грубый, но, хочется надеяться, поучительный расчет верхней границы вероятности возникновения совместной репликационно-трансляционной системы в наблюдаемой части Вселенной; эта вероятность, несомненно, исчезающе мала. Противоположное предсказание состоит в том, что любые формы жизни, которые могут быть обнаружены на Марсе, или, возможно, Европе (спутнике Юпитера, на котором была обнаружена жидкая вода), или даже на любой экзопланете в ходе будущих планетарных исследований, будут иметь общее с земной жизнью происхождение. Любая из этих находок опровергнет сильную гипотезу АХЭ, но не сделает модель МММ незначимой для нашего понимания происхождения жизни. Действительно, любое такое открытие (важное само по себе) просто понизит порог биологической эволюции в масштабе рис. 12-7.
Читать дальшеИнтервал:
Закладка:








