Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Эволюция в мире микробов и вирусов и трехдоменное древо жизни
Вероятно, в ходе развития биологии наибольшее влияние на изменение представления об эволюции оказало распространение эволюционных исследований на мир микробов, а именно одноклеточных эукариот (протист), прокариот (бактерий и архей) и вирусов. Дарвиновское представление об эволюции и все достижения эволюционной биологии нескольких последующих десятилетий базировались исключительно на исследовании животных и растений, тогда как одноклеточные эукариоты (протисты) и бактерии (монеры) были сугубо номинально размещены у корня древа жизни Эрнстом Геккелем и его последователями. Хотя к 1950-м годам генетический анализ бактериофагов и бактерий продвинулся настолько, что стало очевидным, что эти формы жизни обладают эволюционирующими геномами, СТЭ не принимала во внимание эти открытия. То, что бактерии (не говоря уже о вирусах) эволюционируют по тому же самому принципу и с использованием тех же механизмов, что и животные и растения, отнюдь не очевидно, учитывая все их разительные биологические отличия от многоклеточных организмов, и в особенности из-за отсутствия у них типичного полового размножения и репродуктивной изоляции, ключевых для видообразования среди животных и растений.
Фактически прокариоты стали «видны» эволюционным биологам в 1977 году, после выхода революционной работы Вёзе и его коллег по филогенезу рРНК (Woese, 1987) [26]. Рассмотренное в общем контексте, открытие Вёзе является эпохально важным и, возможно, даже заслуживает сравнения с открытием структуры ДНК. Вёзе установил, что в одной молекулярной структуре, а именно последовательности нуклеотидов рРНК, выявляется очевидная консервативность во всем диапазоне клеточных форм жизни. Кроме того, чрезвычайно информативным оказался и филогенетический анализ этой универсальной консервативной молекулы: он показал, что рРНК, в некотором приближении, эволюционирует с постоянной скоростью, то есть подчиняется модели молекулярных часов. Это привело к еще одному важному открытию, ставшему одним из символов эволюционной биологии конца ХХ века, – трехдоменному древу жизни (см. рис. 2–3; Woese et al., 1990). Тремя доменами являются бактерии, археи и эукариоты. Домен архей был открыт Джорджем Фоксом и Вёзе сравнительным анализом рРНК, когда в новой группе ничем, казалось бы, не примечательных «бактерий» обнаружились существенные отличия как от остальных бактерий, так и от более сложных эукариотических организмов. В дополнение к разграничению трех доменов, Вёзе и его коллеги использовали филогенетический анализ рРНК для идентификации нескольких основных ветвей архей и бактерий (Woese, 1987). Из этого следовало, что эволюция прокариот столь же доступна для изучения, как эволюция сложных эукариот, – концепция, чуждая микробиологам до работы Вёзе (Stanier and Van Niel, 1962). Благодаря достижениям Вёзе, его сотрудников и последователей появилась все усиливающаяся тенденция приравнивать филогенетическое древо рРНК, с его трехдоменной структурой, к древу жизни Дарвина и Геккеля (Pace, 2009а, 2006). В течение нескольких лет после публикации открытий Вёзе стало ясно, что топологически древо рРНК (по крайней мере, в своих основных чертах) конгруэнтно деревьям некоторых из самых консервативных белков, таких как рибосомные белки, факторы трансляции, субъ единицы ДНК-зависимой РНК-полимеразы и мембранные АТФазы.
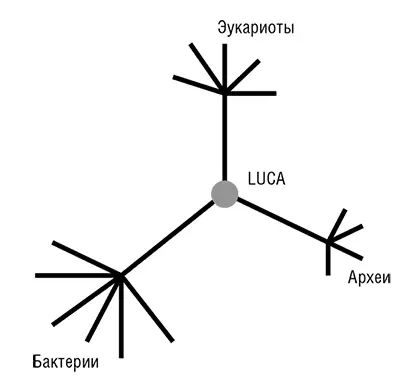
Рис. 2–3. Трехдоменное древо жизни Вёзе.
Две группы исследователей независимо друг от друга пришли к блестящей идее о том, как определить положение корня в эволюционном дереве, которое до этого было бескорневым (рис. 2–3). Для этой цели можно использовать древние паралоги, которые представлены в (почти) всех организмах и, таким образом, можно с уверенностью заключить, возникли в результате дупликации, предшествующей последнему общему предку всех живых организмов (LUCA). Когда дерево строится совместно для двух паралогичных множеств древних ортологов, положение корня между ними определено однозначно, и таким образом корень может быть выведен для каждого из множеств ортологов (см. рис. 2–4; Gogarten et al., 1989; Iwabe et al., 1989). Результаты анализа двух пар древних паралогов, факторов трансляции и субъединиц мембранных АТФаз были полностью совместимы и поместили корень на бактериальную ветвь, установив таким образом кладу архей-эукариотов (см. рис. 2–4). Тем не менее даже в догеномную эпоху было ясно, что не все деревья белок-кодирующих генов имеют ту же топологию, что и дерево рРНК; причины этих отличий оставались неясными и, как предполагалось, включали (за исключением артефактов метода) горизонтальный перенос генов (ГПГ. Smith et al., 1992). Эти расхождения оставались лишь интересным дополнением к трехдоменному ДЖ, но все резко изменилось с наступлением эры геномики.
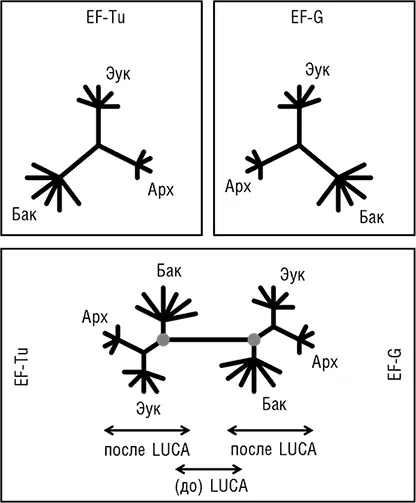
Рис. 2–4. Определение положения корня в трехдоменном древе жизни с помощью древних паралогов. Схематически показаны филогенетические деревья двух широко распространенных факторов инициации трансляции EF-Tu и EF-G, реконструированные независимо (верхние диаграммы) и совместно (нижняя диаграмма). Кружками обозначено вычисленное положение корня в каждом из двух деревьев.
Вирусы и рождение эволюционной геномики
Эволюционная геномика родилась более чем за десять лет до исторического заявления о секвенировании первого бактериального генома. С меньшей помпой (но и не в безвестности) было секвенировано несколько небольших (в рамках 4—100 Кб) различных вирусных геномов, и были разработаны принципы сравнения геномов, наряду с практическими вычислительными методами. Вирусы являются облигатными внутриклеточными паразитами, и вирусные геномы намного меньше и качественно отличаются от геномов клеточных форм жизни. Вирусы обычно не лишены некоторых классов генов, вездесущих и незаменимых в клеточных организмах, таких как гены компонентов системы трансляции и биогенеза мембран. Тем не менее вирусы следуют своей собственной «биологической стратегии» и кодируют все субъединицы вириона, а также по крайней мере некоторые белки, участвующие в репликации вирусного генома. (Одна из центральных тем этой книги – ключевая роль вирусов в эволюции биосферы, так что я возвращаюсь к ней достаточно подробно в гл. 10 [27].) Несмотря на быструю эволюцию последовательности генома, характерную для вирусов (в особенности вирусов с РНК-геномом), эти ранние сравнительные геномные исследования успешно выявили множества генов, консервативных в больших группах вирусов (Koonin and Dolja, 1993). Возможность структурного и функционального картирования всего генома определенной формы жизни была реализована в этих исследованиях впервые, и это стало краеугольным камнем эволюционной геномики. Кроме того, было сделано непредвиденное и важное обобщение: в то время как некоторые гены консервативны для удивительно обширного разнообразия вирусов, архитектура генома, структура вириона и биологические свойства вирусов демонстрируют гораздо большую пластичность (см. гл. 5 и 10, где этот вопрос обсуждается подробнее).
Читать дальшеИнтервал:
Закладка:








