Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Молекулярная датировка событий эволюционного расхождения – весьма специализированная область исследований с множеством сложных технических проблем (Bromham and Penny, 2003; Graur and Martin, 2004). Нам придется пропустить основную часть технических подробностей и перейти прямо к результатам. Заметим что принцип состоит в том, чтобы сопоставить молекулярные данные о дивергенции (то есть результаты сравнения последовательностей, привязанные к филогенетическому древу) с палеонтологическими свидетельствами, используя нескольких точно датированных ископаемых в качестве калибровочных точек (например, самые ранние несомненные ископаемые млекопитающие датируются примерно 120 миллионами лет назад, так что это – самое позднее время начала распространения млекопитающих). Принимая строгие или нестрогие молекулярные часы, можно получить временную оценку для любого события дивергенции относительно взятых калибровочных точек и для данной топологии древа. Такие оценки могут быть достаточно достоверны, когда они представляют собой интерполяцию (то есть вывод о времени дивергенции внутри временного интервала, ограниченного калибровочными точками), но гораздо менее надежны, если получены путем экстраполяции (даты за пределами калибровочного интервала). К сожалению, для древних датировок, таких как возраст LECA, экстраполяция неизбежна. Временные оценки, полученные разными исследователями, охватывают чрезвычайно широкий диапазон дат между 1000 миллионами лет назад и 2300 миллионами лет назад. Несколько недавних, независимых и технически продвинутых исследований, использовавших нестрогие модели молекулярных часов или РГИ с похожим поведением в ходе эволюции, независимо привели к концепции «молодого LECA», которая помещает первичное расхождение эукариот в промежуток примерно от 1100 до 1300 миллионов лет назад (Chernikova et al., 2011; Douzery et al., 2004). Конечно, проблема не решена, но это, по-видимому, наилучшая из имеющихся на сегодня оценок возраста LECA. Из такой оценки следует долгая стволовая фаза в несколько сотен миллионов лет (см. рис. 7–3 б ), поскольку не вызывающие сомнений окаменелости эукариот относятся ко времени более чем 1500 миллионов лет назад (Knoll et al., 2006).
Этот вывод заставляет серьезно переоценить наши современные знания о ранней эволюции эукариот. С одной стороны, результаты реконструкции, описывающие LECA как одноклеточного эукариота современного типа с полностью развитыми характерными функциональными системами эукариотической клетки, становятся менее неожиданными: действительно, похоже, что имелось достаточно времени для эволюции этих черт с момента появления (примитивной) эукариотической клетки. То же самое верно и для ряда генных дупликаций, отнесенных к LECA: согласно сценарию длинного ствола, они не произошли взрывообразно; было достаточно времени, чтобы гены дуплицировались постепенно. С другой стороны, стволовая фаза – это настоящие темные века в эволюции эукариот: мы не знаем о них почти ничего и можем надеяться узнать предельно мало. В самом деле, LECA по сути является «горизонтом событий» для сравнительной геномики: используя только сравнение геномов, мы не можем заглянуть в фазу ствола. Можно получить некоторое представление о том, что тогда происходило, тщательно изучая дупликации предковых эукариотических генов, но это практически единственный источник информации о стволовой фазе. Мы не имеем представления о реальном разнообразии эукариот темных веков, и надежда на то, что мы сможем оценить его в будущем, очень мала. Данные ископаемых выявляют некоторое разнообразие, но эта летопись никогда не будет полной, и трудно даже сказать, насколько она неполна. За немногими исключениям, ископаемые останки эукариот раннего и среднего протерозоя вряд ли представляют какой-нибудь из существующих ныне таксонов – наблюдение, которое следует интерпретировать осторожно, но которое в принципе согласуется со сценарием «молодого LECA» и идеей о существовании некоторого вымершего разнообразия, которое в настоящее время недоступно для наблюдения (см. рис. 7–3 б ). Очевидна возможность, что LECA был тем организмом-«первооткрывателем», который захватил митохондриального эндосимбионта, и что эндосимбиоз инициировал распространение дошедших до нас эукариот. Из этого сценария следует, что предки LECA, разнообразные эукариотические Protozoa , представляют вымершую, первично амитохондриальную биоту Archezoa . Однако есть другая, возможно более убедительная версия – что митохондриальный эндосимбиоз на самом деле дал начало появлению эукариот как таковому, так что эукариоты темных веков уже содержали митохондрии либо МПО. Мы обсудим эту дилемму и доводы в поддержку гипотезы об инициированном эндосимбиозом возникновении эукариот ниже в этой главе, после того как рассмотрим связи между эукариотами и прокариотами, обнаруженные путем сравнения геномов.
Корни эукариот среди архей и бактерий
Поиск архейного и бактериального «родителей» эукариот
Все эукариоты являются гибридными (химерными) организмами как в смысле клеточной организации, так и в отношении набора генов. Действительно, как отмечалось ранее в этой главе, все ныне существующие эукариоты, по-видимому, обладают митохондриями либо МПО, произошедшими от альфа-протеобактерий, а растения и многие группы Chromalveolata к тому же содержат пластиды, произошедшие от цианобактерий. Набор генов у эукариот представляет собой разнородную смесь генов, вероятно происходящих от архей, генов с наиболее вероятным бактериальным происхождением и генов неизвестного происхождения, считающихся в настоящее время специфическими для эукариот. Может показаться парадоксальным, что, хотя филогенетические деревья, основанные на генах рРНК и соединенных последовательностях белков, участвующих в передаче информации, таких как полимеразы, рибосомальные белки и субъединицы сплайсосом, уверенно объединяют архей и эукариот, независимые сравнения полных геномов сходятся на том, что у эукариот в три и более раза больше генов с ближайшими бактериальными гомологами, чем с ближайшими архейными гомологами (см. рис. 7–4; Esser et al., 2004; Koonin et al., 2004; Makarova et al., 2005). Архейный набор значительно обогащен генами, связанными с функциями обработки информации (трансляция, транскрипция, репликация, сплайсинг), тогда как бактериальный набор кодирует в основном ферменты метаболизма, мембранные белки и компоненты биогенеза мембран, различные сигнальные молекулы и другие «операциональные» белки (подробнее см. ниже в этой главе).
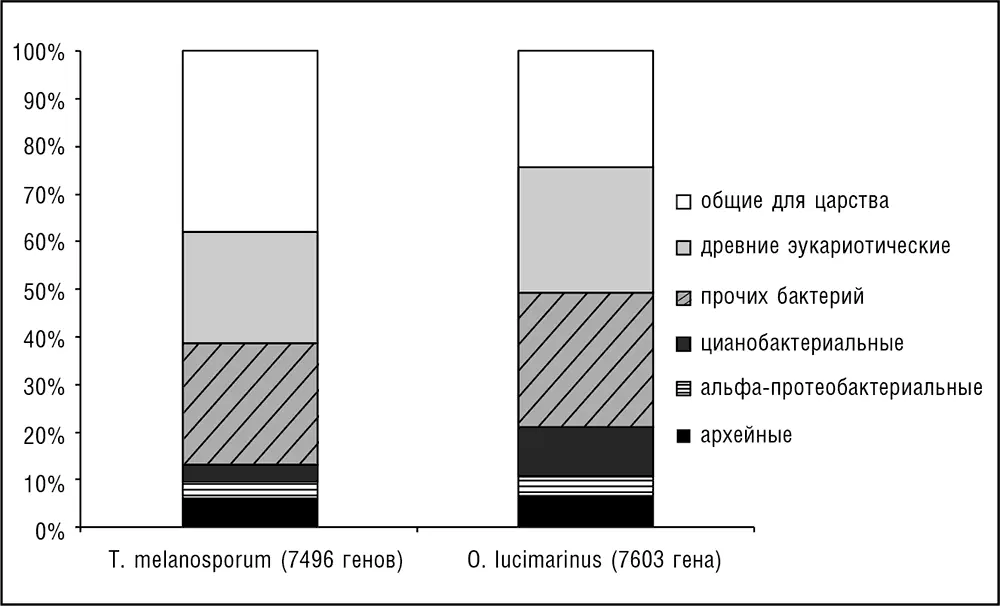
Рис. 7–4. Распределение генов двух дальнородственных эукариот, согласно их предполагаемому происхождению: архейные, бактериальные либо специфичные для эукариот. Последовательности всех кодируемых белков гриба Tuber melanosporum (черный трюфель) и зеленой водоросли Ostreococcus lucimarinus сравнивались с базой данных NCBI RefSeq программой BLASTP (Altschul et al., 1997), а предполагаемое филогенетическое родство для каждого белок-кодирующего гена определялось с использованием специально написанной программы. Стоит отметить похожие, относительно небольшие фракции генов очевидно альфа-протеобактериального происхождения и более крупную фракцию цианобактериальных генов у водоросли.
Читать дальшеИнтервал:
Закладка:








