Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
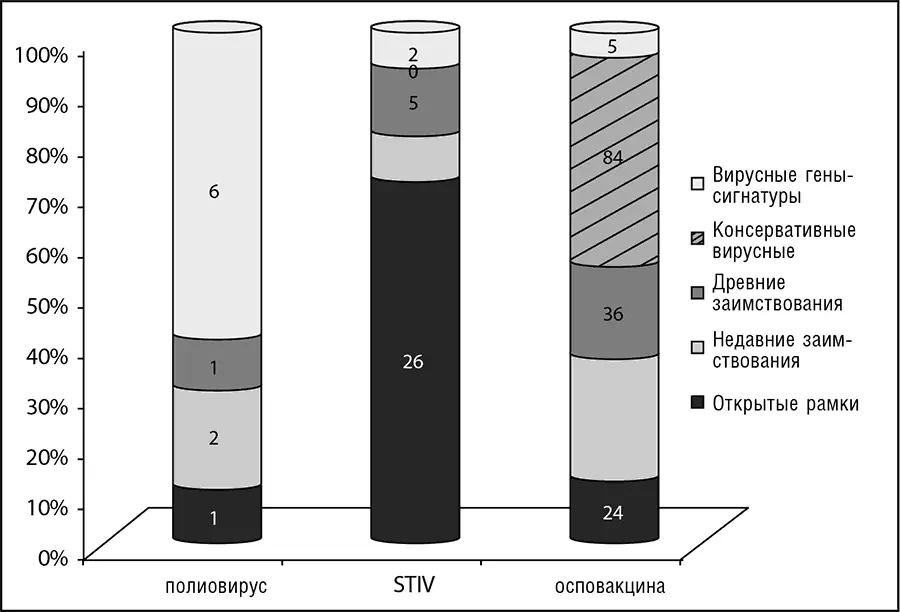
Рис. 10-2. Разбиение вирусных генов на пять эволюционных классов: вирус, обладающий малым геномом: полиовирус (7,4 Кб); вирус, обладающий геномом промежуточного размера: Sulfolobus turreted icosahedral virus (STIV); вирус, обладающий крупным геномом: вирус осповакцины (195 Кб). Данные по Koonin et.al., 2006.
Другие белки, перечисленные в табл. 10-2, не столь широко распространены, как белок капсида с укладкой типа рулета и геликаза суперсемейства 3, но все же за их счет формируются множественные непредвиденные связи между группами вирусов, во всех других отношениях казавшихся неродственными. В качестве примера можно рассмотреть эндонуклеазу инициации репликации по механизму «катящегося кольца», которая объединяет огромное разнообразие небольших репликонов одноцепочечных и двухцепочечных ДНК, включая вирусы, плазмиды и транспозоны, которые размножаются в животных, растениях, бактериях и археях. Детальный анализ аминокислотных последовательностей показал, что ДНК-связывающий домен репликативного белка вирусов полиомы и папилломы (например, T -антигена SV40) представляет собой неактивную производную форму эндонуклеазы инициации репликации способом «катящегося кольца» (Iyer et al., 2005). Таким образом, посредством детального исследования одного из белков-сигнатур хорошо известная связь между различными небольшими репликонами оцДНК (вирусов и плазмид) распространилась также на группу репликонов дцДНК такого же размера. Подобное расширение набора вирусных групп, охватываемых определенным геном-сигнатурой, произошло также в результате детального анализа АТФазы, отвечающей за упаковку вирусной ДНК в капсид, и праймазы архей и эукариот, участвующей в инициации репликации ДНК (Iyer et al., 2005; Iyer et al., 2004b; см. табл. 10-2).
Репликация генома РНК-вирусов с позитивным геномом, дцРНК-вирусов, вирусов с негативным геномом и обратно транскрибирующихся вирусов (элементов) катализируется другим классом ферментов – вирусных сигнатур: РНК-зависимыми РНК-полимеразами и обратной транскриптазой. Полимеразы РНК-вирусов с позитивным геномом и обратная транскриптаза образуют монофилетическую группу внутри обширного класса так называемых palm-доменов («домены-ладони»), характерных для различных полимераз (Iyer et al., 2005; Koonin et al., 2008). РНК-зависимые РНК-полимеразы дцРНК-вирусов и вирусов с негативным геномом, скорее всего, сильно измененные производные того же полимеразного домена (Delarue et al., 1990; Gorbalenya et al., 2002; Koonin et al., 1989). Этот ген-сигнатура может привести нас к самым ранним этапам эволюции жизни, к миру РНК (см. гл. 11 и 12 – там гораздо подробней) и началу мира вирусов. Palm-домен, вероятно, является изначальным белком-полимеразой, сменившим рибозимные полимеразы (гипотетического) мира РНК. Это предположение поддерживается не только широким распространением palm-домена среди современных форм жизни, но еще и структурной, а следовательно, и эволюционной связью между palm-доменом и доменом, содержащим РНК-распознающий мотив (RRM, RNA Recognition Motif), древний РНК-связывающий домен, который первоначально, возможно, способствовал репликации рибозимов (Aravind et al., 2002). РНК-зависимые РНК-полимеразы и обратные транскриптазы исключены из основного хода репликативного цикла клеточных форм жизни, хотя большинство эукариотических геномов, особенно растений и животных, включает в себя множество копий RT-кодирующих ретроэлементов; у прокариот тоже есть немного подобных элементов (см. также гл. 5 и 7). Эти элементы, однако, эгоистичны и с эволюционной точки зрения принадлежат миру вирусов. Возможно, наиболее впечатляющее вторжение обратной транскриптазы в мир клеток – это каталитическая субъединица эукариотической теломеразы, важнейшего фермента, участвующего в репликации концов хромосом [107]. Конечно, не стоит забывать, что все интроны эукариот произошли от ретроэлементов прокариот (см. гл. 7). Примечательно, что единственная другая известная РНК-зависимая РНК-полимераза, не родственная полимеразам, содержащим palm-домен, и являющаяся компонентом системы РНК-интерференции эукариот (см. гл. 7), также, по-видимому, имеет вирусное происхождение (Iyer et al., 2003).
Таблица 10-2. Белки, кодируемые наиболее распространенными генами – вирусными сигнатурами.
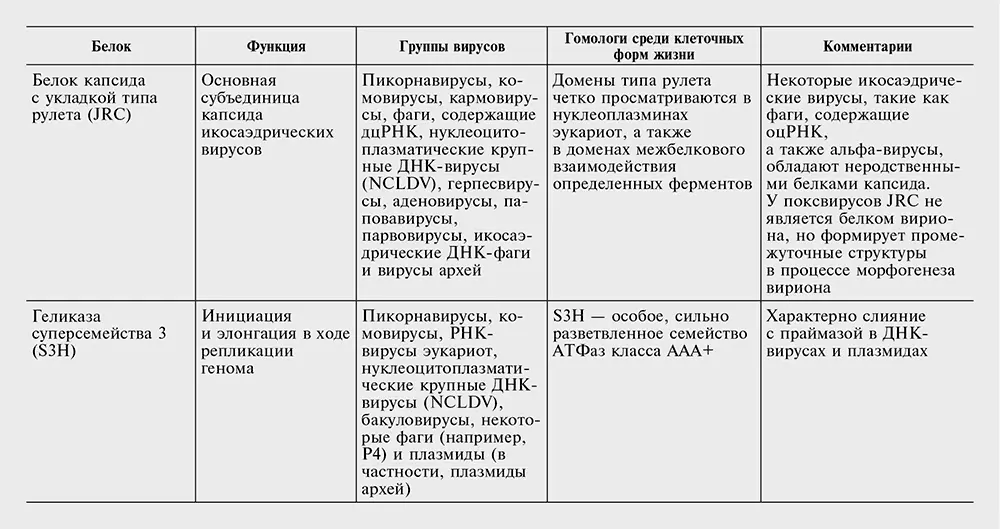
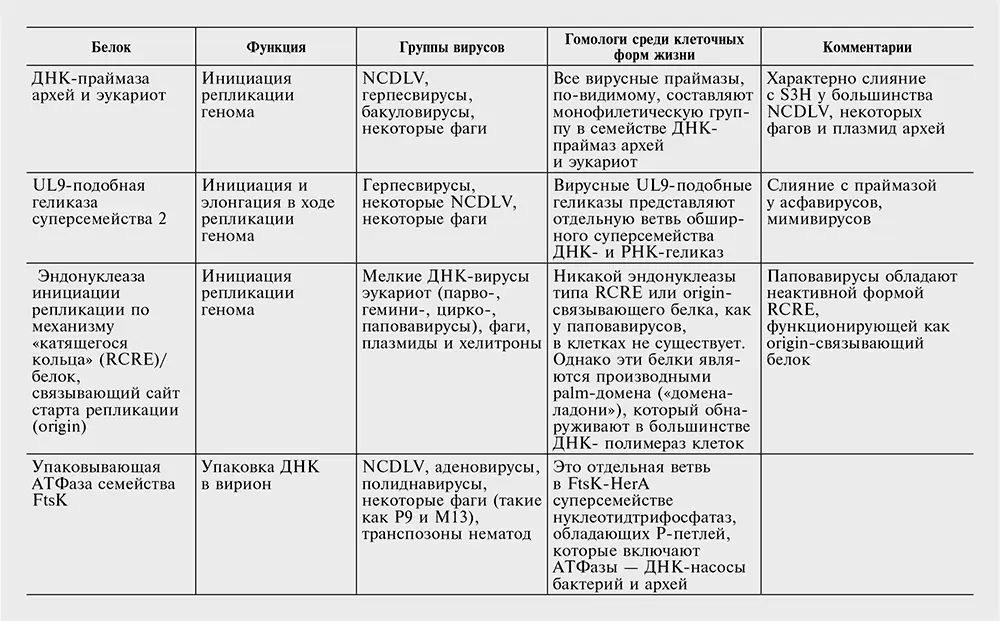
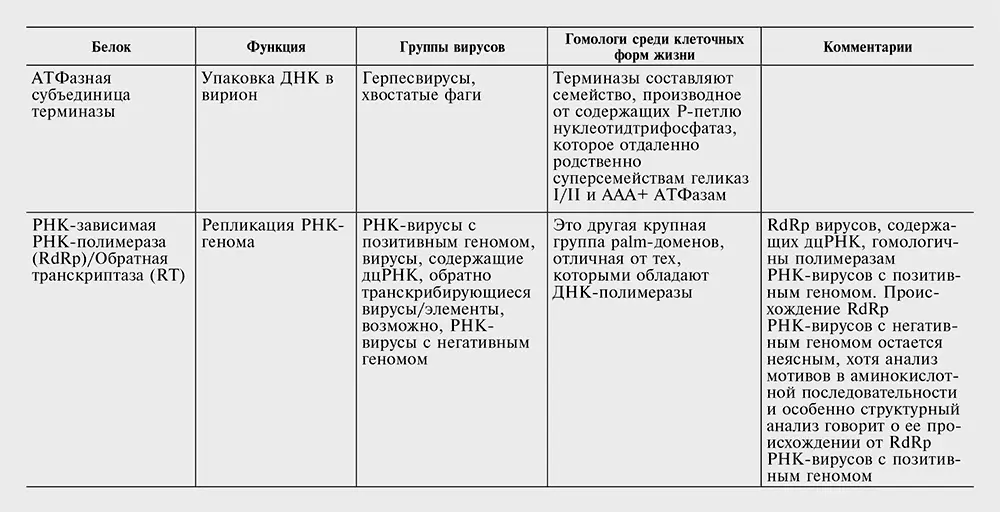
Перечень генов – вирусных сигнатур в табл. 10-2 консервативен. Вероятнее всего, другие гены также заслуживают статуса сигнатур, но отыскать явные свидетельства в пользу этого непросто. Секвенирование новых вирусных геномов в совокупности с всесторонним сравнительным анализом могло бы помочь выявить новые гены, которые, несмотря на относительно узкое распространение среди вирусов, могут считаться «сигнатурами». В самом деле, так может обстоять дело со многими, если не с большинством, генов класса 4, вирусных генов, консервативных для больших групп вирусов, но не клеточных форм жизни.
Комбинация свойств белков – вирусных сигнатур во многом необычна и требует эволюционистского объяснения. В самом деле, все гены-сигнатуры без исключения ответственны за важнейшие, центральные аспекты вирусного жизненного цикла, включая репликацию генома, формирование вириона и упаковку геномной ДНК в вирион (см. табл. 10-2). Обладание этими генами связывает между собой совершенно различные классы вирусов, которые часто имеют совершенно разные стратегии самовоспроизведения и различаются по размеру генома на три порядка. Наконец, у всех генов – вирусных сигнатур есть отдаленные гомологи в клеточных формах жизни (см. табл. 10-2), но вирусные версии, по-видимому, имеют общее происхождение.
Две сразу возникающие гипотезы о происхождении вирусных генов-сигнатур предлагают противоречащие друг другу эволюционные сценарии, ответственные за существование и распространение этих генов (Koonin et al., 2006).
1. Гены-сигнатуры – наследие последнего универсального общего предка вирусов (Last Universal Common Ancestor of Viruses , LUCAV). Этот сценарий предполагает, что, невзирая на все свидетельства в пользу противоположного (см. выше), все ныне живущие вирусы на самом деле монофилетичны, хотя их дальнейшая эволюция включала в себя масштабную потерю генов в некоторых эволюционных линиях, а также обильное заимствование новых генов от хозяев в других.
2. Напротив, в рамках гипотезы о полифилетическом происхождении вирусов распространение генов-сигнатур по всему диапазону групп вирусов может объяснить горизонтальный перенос генов.
Читать дальшеИнтервал:
Закладка:








