Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
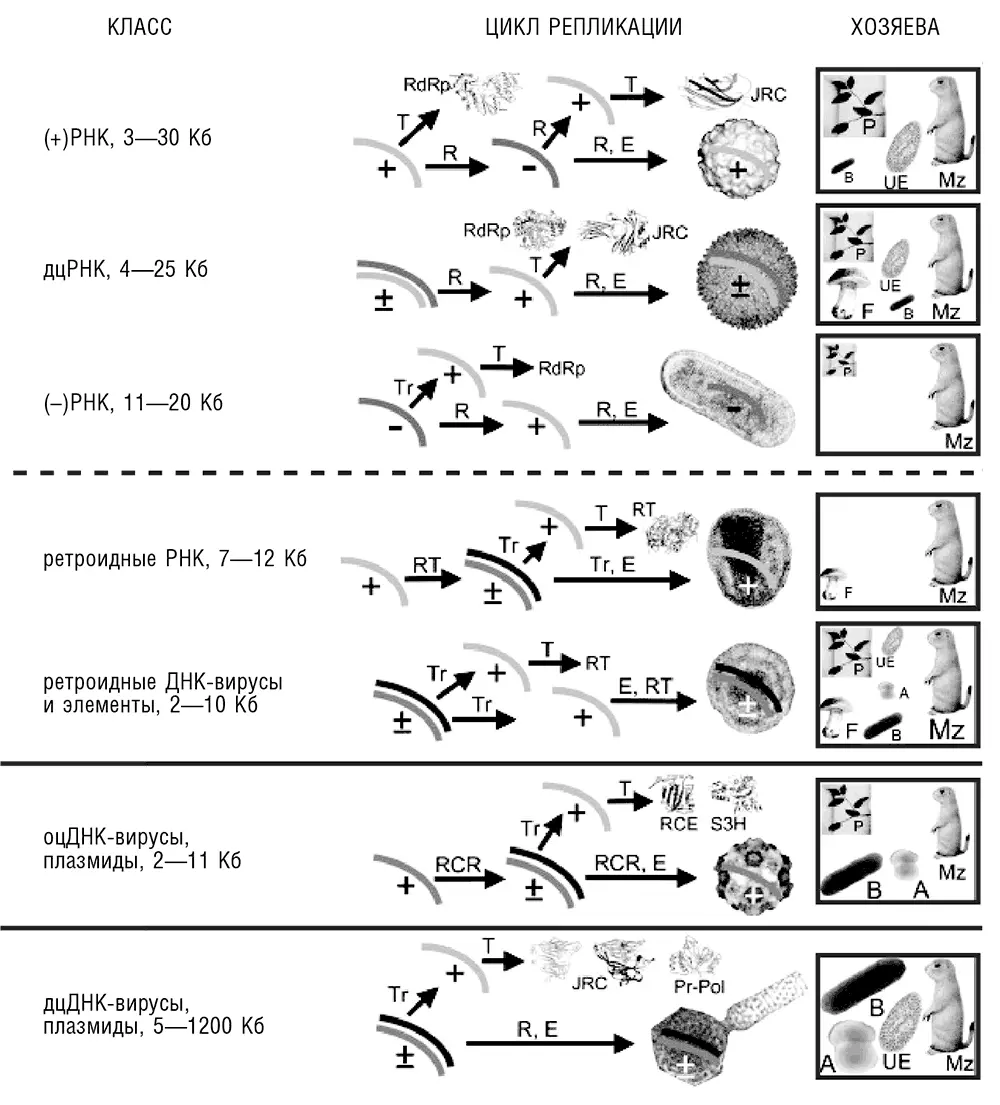
Рис. 10-1. Разнообразие стратегий репликации-экспрессии у вирусов и вирусоподобных элементов. Для каждого класса вирусов и схожих элементов указаны приблизительные размеры генома (Кб, в тысячах оснований). «+» обозначает плюс-цепь (однонаправленную с мРНК), «—» обозначает минус-цепь. Tr– транскрипция, T – трансляция, R – репликация, E – капсидирование вируса, A – археи, B – бактерии, F – грибы, Mz – животные, P – растения, UE – одноклеточные эукариоты. Для каждого класса вирусов (элементов) показаны типичные структуры их белков-маркеров, а также ультрамикроскопические изображения характерных представителей. RdRp – РНК-зависимая РНК-полимераза; JRC – белок капсида с укладкой типа рулета; RT – обратная транскриптаза; RCR – (инициирующая) эндонуклеаза репликации способом катящегося кольца. Справа показан диапазон возможных хозяев. Размер соответствующего изображения и сокращения приблизительно соответствует представленности данного класса вирусов в этом таксоне (по Koonin et al., 2006, с изменениями).
Диапазон сложности геномов, их функционального содержания и разнообразие геномной архитектуры вирусов
Диапазон размера геномов вирусов с различными геномными стратегиями весьма велик: размеры геномов самого крупного известного вируса – мимивируса – и самых мелких вирусов (таких как цирковирусы) отличаются на три порядка. Если мы включим сюда вироиды, которые не кодируют белки, но являются полноценными эгоистичными генетическими элементами и даже патогенами, размах колебаний расширяется до четырех порядков (см. рис. 10-1). Если иметь в виду, что в геномах вирусов обычно белок-кодирующие гены расположены вплотную, то колебания количества генов укладываются приблизительно в тот же диапазон. Размер генома сильно зависит от природы генома и цикла репликации-экспрессии. Видимо, лишь у вирусов, содержащих дцДНК, геном может достичь больших (по вирусным меркам) размеров, более 35 Кб, и (на данный момент) вплоть до 1,1 Мб (Van Etten et al., 2010). Все классы РНК-вирусов, все элементы, способные к обратной транскрипции, и все вирусы, содержащие оцДНК, обладают небольшими геномами, никогда не превышающими 35 Кб, – и то к этому пределу приближается лишь одна группа довольно редких РНК-вирусов животных (коронавирусы и родственные им формы, составляющие порядок Nidovirales (Gorbalenya et al., 2006). Причина этого очевидна: бульшая химическая стабильность и регулярная структура дцДНК благоприятна для функций хранения информации и репликации, а далее эволюция систем репарации дцДНК еще больше усилила функциональное разделение между дцДНК и другими формами нуклеиновых кислот. В дополнение к разнообразию стратегий репликации-экспрессии и широкому диапазону размеров, вирусные геномы принимают все возможные молекулярные конфигурации, включая как линейные, так и циркулярные молекулы ДНК или РНК и как единичные, так и разделенные на множественные сегменты геномы (хромосомы, см. рис. 10-1).
Функциональный набор вирусных генов разительно различается в зависимости от цикла экспрессии и, что еще важнее, от размера генома и генетической сложности. Малые вирусные геномы кодируют практически исключительно белки, непосредственно участвующие в репликации генома, а также субъединицы вириона. Часто – и для всех известных РНК-вирусов и элементов, способных к обратной транскрипции, это именно так, – вирус кодирует полимеразу, участвующую в репликации его собственного генома. Это легко объяснить: клеточные хозяева вируса обычно не кодируют РНК-зависимой РНК-полимеразы или обратной транскриптазы, которая могла бы осуществить репликацию или обратную транскрипцию длинных молекул РНК. РНК-зависимые РНК-полимеразы и обратные транскриптазы, которые кодируются в геномах клеточных форм жизни и осуществляют «обычные» клеточные функции, такие как теломераза и РНК-зависимая РНК-полимераза, участвующая в РНК-интерференции эукариот, способны синтезировать лишь короткие олигонуклеотиды (см. гл. 7 и обсуждение ниже в этой главе). У ДНК-вирусов, напротив, есть возможность использовать аппарат репликации (и транскрипции) хозяина, и они широко пользуются этой возможностью. Так, многие вирусы этого типа, в частности большинство известных вирусов, поражающих архей, так же как и многочисленные умеренные бактериофаги (такие как фаг лямбда, классический модельный организм), не кодируют даже репликативной полимеразы и никаких других белков, непосредственно участвующих в репликации. В таких случаях последовательности ДНК, ответственные за распознавание и привлечение репликативного аппарата хозяина, оказываются основными факторами, определяющими возможность автономной репродукции вируса (см. определение выше), хотя вирусные белки выполняют другие важные функции в вирусной репродукции, такие как подавление или репрограммирование экспрессии генов и метаболизма хозяина. Вирусы с самыми большими геномами, напротив, вдобавок к белкам, составляющим аппарат репликации вирусного генома, кодируют целую коллекцию разнообразных белков, участвующих в процессах репарации, мембранного транспорта, ряде метаболических путей, а в некоторых случаях и трансляции. Обычно (и во всех случаях, когда затрагивается трансляция) вирус кодирует не систему или путь целиком, а только ферменты для одного-двух шагов, которые дополняют или видоизменяют соответствующие функциональные системы клетки хозяина.
Вирусы, содержащие различные формы нуклеиновых кислот, распределены по таксонам хозяев не равномерным и не случайным образом. В частности, необычайное разнообразие содержащих дцДНК бактериофагов и вирусов архей находится в разительном контрасте с отсутствием дцДНК-вирусов у растений. РНК-вирусы, напротив, чрезвычайно распространены и разнообразны у растений и животных, но среди бактерий на данный момент представлены лишь двумя небольшими семействами и до сих пор не обнаружены у архей [103](см. рис. 10-1). В некоторых случаях биологическая подоплека характерного спектра хозяев вируса совершенно ясна. Например, в растениях крупные вирусы столкнулись бы с серьезными проблемами с межклеточным распространением, так как плазмодесмы (каналы, соединяющие окруженные клеточной стенкой клетки растений) непроницаемы для больших частиц и даже больших молекул ДНК. Однако в большинстве случаев причины предпочтительного распределения вирусов среди тех или иных групп хозяев остаются неизвестными. Например, сложно сказать, почему РНК-вирусы столь распространены среди растений и животных, но не среди прокариот; далее в этой главе мы вернемся к рассмотрению этого вопроса с другой, эволюционной позиции.
Читать дальшеИнтервал:
Закладка:








