Михаил Супотницкий - Микроорганизмы, токсины и эпидемии

- Название:Микроорганизмы, токсины и эпидемии
- Автор:
- Жанр:
- Издательство:Вузовская книга
- Год:2006
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Супотницкий - Микроорганизмы, токсины и эпидемии краткое содержание
Кроме того, в монографии, впервые для отечественной литературы, обстоятельно освещены эпидемиология биологического террористического акта и методология выявления такого преступления. Книга хорошо иллюстрирована, содержит приложения и рассчитана на широкий круг специалистов.
Микроорганизмы, токсины и эпидемии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Патогенность облигатных паразитов (возбудители кори, коклюша, гриппа, желтой лихорадка, чумы, натуральной оспы, лихорадки Денге, Ку-лихорадки и др.). Патогенность таких паразитов является исключительно вынужденной и существует через необходимость сохранения хозяина живым для последующего переноса к новым хозяевам (рис. 4). В зависимости от способа переноса (основные — трансмиссивный и воздушно-капельный), плотности и чувствительности инфицируемой популяции, вирулентность возбудителя новой болезни может значительно колебаться, однако в его распространении всегда можно отследить эпидемическую цепочку. Клиническая картина болезни может носить характер давно сформировавшейся инфекционной патологии, т. е., иметь четкий клинико-патогенетический синдром и эпидемиологию. Наиболее опасно появление в человеческих популяциях новых возбудителей данного типа, использующих трансмиссию с помощью какого либо переносчика, который уменьшает зависимость паразитического микроорганизма от живого хозяина. Более того, он будет поддерживать отбор высоковирулентных штаммов из-за большой вероятности захвата им возбудителя с кровью больных с тяжелыми формами инфекции. Однако процесс формирования таких форм симбиоза хозяина и паразита носит характер коэволюции. Поэтому в инфицированных популяциях новый возбудитель никогда не достигает максимума своей вирулентности, в чем можно убедиться опытным путем, уменьшая его LD 50последовательными пассажами через хозяев. Эффект пассажей делает паразитическую трансмиссию независящей от выживания самого хозяина. Посредством этого ему «дозволяется» большая вирулентность без компромисса, необходимого для его способности передаваться другим хозяевам [Levin В., 1996].
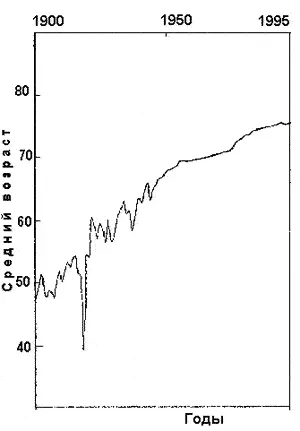
Рис. 4. Снижение средней продолжительности жизни людей на планете после пандемии испанки в 1918 г. и с начала 1960-х гг. Масштабные эпидемии и пандемии, вызываемые облигатными микроорганизмами, использующими первую стратегию паразитизма, способны приводить к опустошению отдельных стран. Патогенность таких паразитов существует через необходимость оставлять хозяина живым. Поэтому численность населения не только восстанавливается, но и может превысить «доэпидемический» период. Однако с начала 1960-х годов рост средней продолжительности жизни замедлился [Lederberg J., 1997]. В XXI столетии возможна ее отрицательная динамика из-за давления, оказываемого возбудителями инфекций, использующими вторую и третью стратегии паразитизма и существующими без необходимости сохранять жизнь своим жертвам
Таким образом, вышеуказанные противоречия «теории коэволюции», созданы теми «коэволюционистами», которые понимают взаимную адаптацию макро- и микроорганизмов, как процесс, ведущий к утрате микроорганизмом паразитических свойств и к превращению его в свою противоположность — комменсал. О.В. Бухарин и Б.Я. Усвяцов (1996), опираясь на предложенный В. Беляковым и соавт. (1987) «принцип саморегуляции паразитарных систем», «развили» эволюционное учение «правилом», в соответствии с которым «эволюция не дает существенных преимуществ ни одному из взаимодействующих видов — она направлена на достижение динамического противоречивого равновесия». Т. е. эволюции как естественного отбора наиболее приспособленных видов, не существует. А эволюция симбиотических систем представлена ими в виде последовательности событий — «комменсализм-паразитизм-аменсализм-комменсализм» и т. д., напоминающей ламарковское упражнение органов. О.В. Бухарин и Б.Я. Усвяцов (1996), так же как и N. Ampel (1991), принимают частный случай снижения вирулентности микроорганизмом в конце эпидемического процесса за общую закономерность — утрату им паразитических свойств в ходе «эволюции симбиотических систем».
Возможна ли вообще утрата паразитических свойств облигатным организмом, как настаивают указанные выше авторы? Чтобы ответить на этот вопрос, посмотрим, какую цену платят другие биологические виды за переход к паразитическому существованию. Например, внутренние паразиты теряют органы чувств, затем у них до предела упрощается нервная система, и, как у ленточных червей, исчезает пищеварительная — она им больше не нужна. Т. е., отбор перестает следить за формированием структуры, и этого достаточно. Остальное делает второе начало термодинамики . Те структуры, которые поддерживает отбор, сохраняются и развиваются, например, органы прикрепления к стенке кишечника [Медников Б.М., 1982]. Эта закономерность носит общебиологический характер. Поэтому она справедлива и для микроорганизмов. Таким образом, переход к облигатному паразитизму всегда сопровождаемся упрощением организма. Организм не может «скачком» вернуть те утраченные структуры, которые бы позволили ему вновь вернуться в прежнюю среду обитания, перейти к другим формам симбиоза, либо изменить стратегию паразитизма. Для этого ему нужно стать другим видом. А так как это менее вероятно, то при смене условий окружающей среды происходит либо его замещение другим паразитом, либо формирование других форм симбиоза, которые могут не включать паразитические микроорганизмы. Следовательно, патогенность облигатных паразитов является адаптивным признаком , а их поддержание в популяциях отдельных видов может достигаться за счет других механизмов (гетерогенность популяции паразита по вирулентности, использование им переносчиков, сохранение в резервуарах и т. п.). Даже сильное отрицательное давление против высокопатогенных паразитов, вызванное небольшой численностью хозяина, либо ограниченным диапазоном паразитизма возбудителя, ведет не к становлению его как комменсала, а к его вытеснению из популяции хозяина новым микроорганизмом с другой стратегией паразитизма (второго и/или третьего типа). Поэтому теории коэволюции не противоречит ни длительная ассоциация патогенных видов микроорганизмов с их хозяевами, ни поддержание патогенности естественным отбором, но только тогда, когда речь идет об облигатных паразитах и исключается придуманная некоторыми исследователями возможность их перехода к другим формам симбиоза.
Патогенность факультативных паразитов(возбудители полиомиелита, холеры, гепатита А, ботулизма, бактериальных менингитов и др.). Для возбудителей инфекционных болезней, являющихся факультативными паразитами, характерны фекально-оральный и воздушно-капельный пути передачи, высокая степень носительства и лишь эпизодическое проявление их в форме тяжелых инфекций. Так, на одного больного менингитом приходится 180 бактерионосителей [Покровский и др., 1976]. По данным В.В. Алексеенко (1991) соотношение больных холерой и вибрионосителей колеблется в разных регионах от 1:1 до 1:100. Проникновение вируса полиомиелита через гематоэнцефалический барьер происходит не чаще чем в 1 % случае от числа инфицированных [Болотовский В.М., 1993]. Колонизация факультативными паразитами слизистых поверхностей макроорганизма (кишечник, носоглотка) осуществляется в условия противодействия со стороны микробов-антагонистов, местных барьерных факторов и др. Поэтому создаются условия для природной селекции высоковирулентных штаммов в виде локального феномена. Это позволяет паразитическому микроорганизму:
Читать дальшеИнтервал:
Закладка:





![Михаил Супотницкий - Очерки истории чумы. Книга II. Чума бактериологического периода [без иллюстраций]](/books/1084186/mihail-supotnickij-ocherki-istorii-chumy-kniga-ii.webp)
![Михаил Супотницкий - Очерки истории чумы. Книга I. Чума добактериологического периода [без иллюстраций]](/books/1084187/mihail-supotnickij-ocherki-istorii-chumy-kniga-i-ch.webp)


