Александр Древаль - Эндокринные синдромы и болезни. Руководство для врачей

- Название:Эндокринные синдромы и болезни. Руководство для врачей
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:2020
- Город:Москва
- ISBN:978-0-36940-352-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Древаль - Эндокринные синдромы и болезни. Руководство для врачей краткое содержание
Пособие облегчит студентам старших курсов медицинских вузов, клиническим ординаторам и начинающим врачам усвоение фундаментальных принципов дифференциальной диагностики. Книга поможет им стать отличными специалистами с высокой общеклинической культурой и эффективно применить знания в реальной клинической практике.
Эндокринные синдромы и болезни. Руководство для врачей - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
3.1.4. Синтез, хранение и секреция гормонов
Пептидные гормоны синтезируются по тому же механизму, что и любые другие белки. Часто сначала синтезируется большая молекула прогормона, которая затем преобразуется в гормон меньших размеров. К примеру, препропаратиреоидный гормон пропаратиреоидный гормон паратиреоидный гормон. С другой стороны, стероиды и катехоламины синтезируются из меньших молекул.
Эндокринные органы не являются уникальным местом синтеза гормонов (к примеру, ХГЧ синтезируется практически во всех тканях организма), вместе с тем только в них синтез гормонов и его регулирование происходят наиболее эффективно. Три основные особенности отличают эндокринный орган от неэндокринной ткани, синтезирующей какой-либо гормон:
• скорость синтеза намного выше в эндокринном органе;
• превращение прогормона в гормон осуществляется только в эндокринном органе (например, проопиомеланокортин превращается в АКТГ в гипофизе, но не в мозге);
• эндокринные железы снабжены механизмом транспорта гормона в кровь, который, как правило, регулируем.
Скорость секреции гормона железой определяется скоростью его синтеза, которую могут регулировать другие, тропные по отношению к этой железе гормоны. За исключением Т 4и 1,25-дигидроксихолекальциферола, запасы гормонов в организме очень ограничены.
Стимуляция секреции гормона связана с деполяризацией клеточной мембраны и открытием кальциевых каналов, что приводит к поступлению кальция в клетку, где он соединяется с кальций-связывающим протеином. Повышение внутриклеточной концентрации кальция необходимо для активации экзоцитоза – процесса, в результате которого содержимое внутриклеточных везикул (гормонов, трансмиттеров) поступает в межклеточное пространство.
3.1.5. Транспорт и элиминация гормонов
Секретируемые в кровь гормоны транспортируются к тканям-мишеням.
• Стероиды и тиреоидные гормоны циркулируют в крови в связанном с белками плазмы состоянии.
• Полипептидные гормоны и катехоламины находятся в крови в свободном состоянии.
На уровне клеток гормоны взаимодействуют со специфическими рецепторами, расположенными:
• на клеточной мембране (кортикотропин, пролактин, ТТГ и ПТГ) – стимулируют образование циклического аденозинмонофосфата (цАМФ), который, в свою очередь, активирует протеинкиназу, участвующую в синтезе белка;
• в цитоплазме (кортизол, тестостерон и эстрадиол) – образуют комплекс с рецептором, который транспортируется в ядро, где связывается с хроматином; это в конечном итоге также изменяет синтез белка.
Гормоны элиминируются из крови в результате метаболических процессов, например пептидные гормоны инактивируются протеолитическими ферментами. В печени гормоны соединяются с глюкуроновой кислотой и секретируются с жёлчью, но при этом частично реабсорбируются, включаясь в так называемый энтеропечёночный цикл. Гормоны также выводятся с мочой.
Небольшие молекулы гормонов (Т 4, в частности) связываются с белками крови, что замедляет их элиминацию из крови и поддерживает небольшой пул свободного гормона в крови на необходимом уровне. Связывание с белками также облегчает транспорт жирорастворимых стероидов.
3.1.6. Гормональные рецепторы
Рецепторы гормонов – клеточные белки, которые связывают гормон. Они находятся на клеточной мембране, в цитоплазме или ядре и реализуют две функции гормонов на уровне клетки:
• селективность;
• взаимодействие с эффекторными механизмами клетки.
Взаимодействие с гормоном вызывает в рецепторе конформационное изменение, что активирует специфическую клеточную ферментную систему, которая фактически и реализует характерный эффект гормона (рис. 3.1). Когда гормон соединяется с рецептором клеточной мембраны, в цитозоле появляются так называемые вторые мессенджеры (первый – гормон), которые, в свою очередь, активируют ферментную систему клетки. В ядре клетки гормон-рецепторный комплекс связан с дезоксирибонуклеиновой кислотой (ДНК) и регулирует экспрессию гена. Максимальный эффект гормона обычно проявляется уже в том случае, если связано менее 50 % рецепторов. Освободившиеся от связи с гормоном свободные рецепторы снова возвращаются в цитозоль или на мембрану клетки, где продолжают участвовать в гормон-рецепторном взаимодействии.
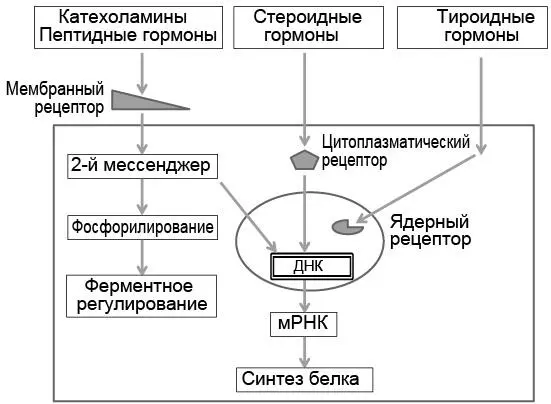
Рис. 3.1.Взаимодействие гормонов с клеточными рецепторами цитоплазматической мембраны, цитоплазмы и ядра. ДНК – дезоксирибонуклеиновая кислота; мРНК – матричная рибонуклеиновая кислота
Стероидные гормоны липофильны, поэтому свободно диффундируют через клеточную мембрану, а затем связываются с цитозольными рецепторными белками. Образовавшийся комплекс «стероидный гормон – рецептор» перемещается в ядро клетки, где он обратимо связывается с геном, изменяет ДНК, что модулирует транскрипцию матричной рибонуклеиновой кислоты (РНК).
Т 3связывается с ядерными рецепторными белками, и комплекс Т 3-рецептор, соединяясь с ДНК, стимулирует образование матричной РНК. Нередко стероидные и тиреоидные гормоны действуют синергично, взаимно усиливая специфические эффекты (потенцируют экспрессию гена).
Количество рецепторов клеточной мембраны и внутриклеточных рецепторов изменяется, меняется и сила их связи с гормоном. Клетки миометрия и молочных желёз содержат окситоциновые рецепторы, число которых возрастает под действием эстрогенов (up-регулирование) и снижается под действием прогестерона (down-регулирование). В миокарде содержатся норадреналиновые рецепторы (β1), число которых и сродство к норадреналину повышается под действием тиреоидных гормонов (Т 3/Т 4).
Водорастворимые гормоны (моноамины, аминокислоты и пептиды) связываются с рецепторами мембраны, которая насыщена липидами, поэтому не позволяет свободно диффундировать водорастворимым гормонам через мембрану. В гормональной реакции клетки водорастворимые гормоны называют первыми мессенджерами. В ответ на их взаимодействие с рецептором внутри клетки активируются так называемые вторые мессенджеры – цАМФ, циклический гуанозинмонофосфат, инозитолтрифосфат, ионы кальция, диацилглицерол и др. Ионы кальция служат очень важным вторым мессенджером. Поток ионов кальция через клеточную мембрану в цитозоль контролируется гормон-рецепторной связью, нервными стимулами или модифицируется другими вторыми мессенджерами.
Концентрация гормонов в большинстве случаев составляет 10–10 моль/л. При этом связывание одной молекулы с мембранным рецептором приводит к образованию 10 000 молекул цАМФ в клетке, и в этом отношении цАМФ выступает в качестве молекулярного усилителя гормонального сигнала (в 10 000 раз!). Фосфодиэстераза разрушает цАМФ, поэтому её ингибиторы – теофиллин и кофеин – действуют синергично с гормонами, у которых цАМФ является вторым мессенджером. цАМФ стимулирует катаболические процессы – липолиз, гликогенолиз (глюкагон), глюконеогенез и кетогенез, секрецию инсулина β-клетками поджелудочной железы.
Читать дальшеИнтервал:
Закладка: