Валерий Шульговский - Основы нейрофизиологии

- Название:Основы нейрофизиологии
- Автор:
- Жанр:
- Издательство:Аспект Пресс
- Год:2000
- Город:М.
- ISBN:ISBN 5-7567-0134-6
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Валерий Шульговский - Основы нейрофизиологии краткое содержание
В книге изложены современные представления о функции клеток и нервной регуляции, а также о комплексной иерархической регуляции основных видов деятельности организма. В основу книги положены курсы лекций, прочитанных автором в течение ряда лет в МГУ им. М. В. Ломоносова и Гуманитарном институте.
Для студентов, аспирантов педагогических и гуманитарных университетов.
Основы нейрофизиологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Самая высокая концентрация серотонина обнаружена в эпифизе (pineal gland). Серотонин в эпифизе превращается в мелатонин, который участвует в пигментации кожи, а также влияет у многих животных на активность женских гонад. Содержание как серотонина, так и мелатонина в эпифизе контролируется циклом свет – темнота через нервную симпатическую систему.
Другую группу медиаторов ЦНС составляют аминокислоты. Уже давно известно, что нервная ткань с ее высоким уровнем метаболизма содержит значительные концентрации целого набора аминокислот (перечислены в порядке убывания): глутаминовой кислоты, глутамина, аспарагиновой кислоты, гамма-аминомасляной кислоты (ГАМК).
Глутамат в нервной ткани образуется преимущественно из глюкозы. У млекопитающих больше всего глутамата содержится в конечном мозге и мозжечке, где его концентрация примерно в 2 раза выше, чем в стволе мозга и спинном мозге. В спинном мозге глутамат распределен неравномерно: в задних рогах он находится в большей концентрации, чем в передних. Глутамат является одним из самых распространенных медиаторов в ЦНС.
Постсинаптические рецепторы к глутамату классифицируются в соответствии с аффинностью (сродством) к трем экзогенным агонистам – квисгулату, каинату и N-метил-D-аспартату (NMDA). Ионные каналы, активируемые квисгулатом и каинатом, подобны каналам, которые управляются никотиновыми рецепторами – они пропускают смесь катионов (Na +и K +). Стимуляция NMDA-рецепторов имеет сложный характер активации: ионный ток, который переносится не только Na +и K +, но также Ca ++при открывании ионного канала рецептора, зависит от потенциала мембраны. Вольтзависимая природа этого канала определяется разной степенью его блокирования ионами Mg ++с учетом уровня мембранного потенциала. При потенциале покоя порядка – 75 мВ ионы Mg ++, которые преимущественно находятся в межклеточной среде, конкурируют с ионами Ca ++и Na +за соответствующие каналы мембраны (рис. 2.17). Вследствие того, что ион Mg ++не может пройти через пору, канал блокируется всякий раз, как попадает туда ион Mg ++. Это приводит к уменьшению времени открытого канала и проводимости мембраны. Если мембрану нейрона деполяризовать, то количество ионов Mg ++, которые закрывают ионный канал, снижается и через канал беспрепятственно могут проходить ионы Ca ++, Na +и K +. При редких стимуляциях (потенциал покоя изменяется мало) глутаматергического рецептора ВПСП возникает преимущественно за счет активации квисгулатных и каинатных рецепторов; вклад NMDA-рецепторов незначителен. При длительной деполяризации мембраны (ритмическая стимуляция) магниевый блок удаляется, и NMDA-каналы начинают проводить ионы Ca ++, Na +и K +. Ионы Ca ++через вторичные посредники могут потенцировать (усиливать) минПСП, что может привести, например, к длительному увеличению синаптической проводимости, сохраняющейся часами и даже сутками.

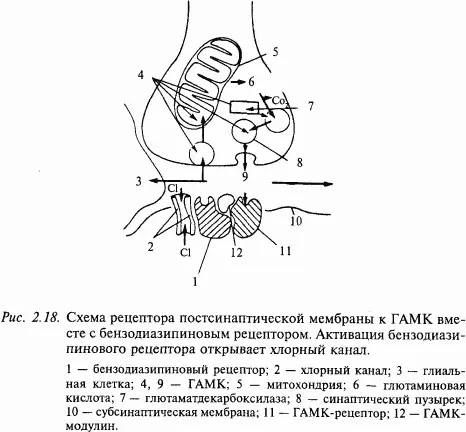
Из тормозных медиаторов ГАМК является самой распространенной в ЦНС. Она синтезируется из L-глутаминовой кислоты в одну стадию ферментом декарбоксилазой, наличие которой является лимитирующим фактором этого медиатора. Известно два типа ГАМК-рецепторов на постсинаптической мембране: ГАМКА (открывает каналы для ионов хлора) и ГАМКБ (открывает в зависимости от типа клетки каналы для. K +или Ca ++). На рис. 2.18 показана схема ГАМК-рецептора. Интересно, что в его состав входит бензодиазипиновый рецептор, наличием которого объясняют действие так называемых малых (дневных) транквилизаторов (седуксена, тазепама и др.). Прекращение действия медиатора в ГАМК-синапсах происходит по принципу обратного всасывания (молекулы медиатора специальным механизмом поглощаются из синаптической щели в цитоплазму нейрона). Из антагонистов ГАМК хорошо известен бикукулин. Он хорошо проходит через гематоэнцефалический барьер, оказывает сильное воздействие на организм даже в малых дозах, вызывая конвульсии и смерть. ГАМК обнаруживается в ряде нейронов мозжечка (в клетках Пуркинье, клетках Гольджи, корзинчатых клетках), гиппокампа (в корзинчатых клетках), в обонятельной луковице и черной субстанции.
Идентификация ГАМК-цепей мозга трудна, так как ГАМК – обычный участник метаболизма в ряде тканей организма. Метаболическая ГАМК не используется как медиатор, хотя в химическом отношении их молекулы одинаковы. ГАМК определяется по ферменту декарбоксилазы. Метод основан на получении у животных антител к декарбоксилазе (антитела экстрагируют, метят и вводят в мозг, где они связываются с декарбоксилазой).
Другим известным тормозным медиатором является глицин. Глицинергические нейроны находятся главным образом в спинном и продолговатом мозге. Считают, что эти клетки выполняют роль тормозных интернейронов.
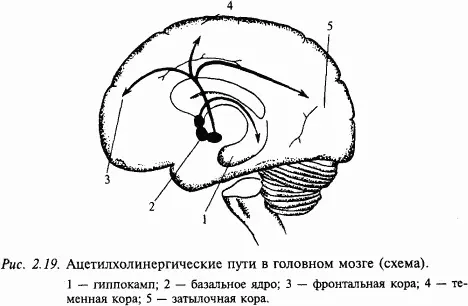
Ацетилхолин – один из первых изученных медиаторов. Он чрезвычайно широко распространен в нервной периферической системе. Примером могут служить мотонейроны спинного мозга и нейроны ядер черепных нервов. Как правило, холинергические цепи в мозге определяют по присутствию фермента холинэстеразы. В головном мозге тела холинергических нейронов находятся в ядре перегородки, ядре диагонального пучка (Брока) и базальных ядрах. Нейроанатомы считают, что эти группы нейронов формируют фактически одну популяцию холинергических нейронов: ядро переднего мозга, nucleus basalis (оно расположено в базальной части переднего мозга) (рис. 2.19). Аксоны соответствующих нейронов проецируются к структурам переднего мозга, особенно в новую кору и гиппокамп. Здесь встречаются оба типа ацетилхолиновых рецепторов (мускариновые и никотиновые), хотя считается, что мускариновые рецепторы доминируют в более рострально распоженных мозговых структурах. По данным последних лет складывается впечатление, что ацетилхолиновая система играет большую роль в процессах, связанных с высшими интегративными функциями, которые требуют участия памяти. Например, показано, что в мозге больных, умерших от болезни Альцгеймера, наблюдается массивная утрата холинергических нейронов в nucleus basalis.
ОПИАТНЫЕ РЕЦЕПТОРЫ И ОПИОИДЫ МОЗГА
Морфин и кодеин – два активных ингредиента морфия. Морфин был очищен в XIX веке и стал широко применяться в медицине. Механизм действия морфина на мозг довольно хорошо изучен благодаря тому, что было синтезировано вещество налоксон, которое является специфическим антагонистом морфина. Сам по себе налоксон неактивен, но действие введенного на его фоне морфина не проявляется. То, что налоксон имеет химическую отруктуру, похожую на морфин, и является его специфическим антагонистом, предполагает наличие на мембране специфических для морфина рецепторов. Использование меченого налоксона показало его специфическое связывание на рецепторах нейронов областей мозга, имеющих отношение к боли. Из мозга голубей было выделено вещество, получившее название энкефалин (т.е. морфин мозга), обладающее свойством морфина. В дальнейшем были открыты другие опиоиды мозга. Эту группу веществ в целом называют эндорфинами.
Читать дальшеИнтервал:
Закладка:









