Александр Коган - Основы физиологии высшей нервной деятельности

- Название:Основы физиологии высшей нервной деятельности
- Автор:
- Жанр:
- Издательство:Высшая школа
- Год:1988
- Город:М.
- ISBN:5-06-001444-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Коган - Основы физиологии высшей нервной деятельности краткое содержание
Учебник состоит из двух частей: общей физиологии высшей нервной деятельности, где изложены общие свойства условных рефлексов и нервных процессов, лежащих в их основе, и частной физиологии высшей нервной деятельности, где рассмотрены особенности условно-рефлекторного поведения основных классов животных, а также высшая нервная деятельность человека. 2-е издание (1-е — 1959 г.) значительно обновлено; особое внимание обращено на возможности использования высшей нервной деятельности для решения практических задач.
Максимальная ширина картинки в этой fb2-книге — 600 пикселей, но большинство картинок имеют размер не более 460 пикселей.
Основы физиологии высшей нервной деятельности - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Анализ свойств ДНК и влияния ингибиторов ее синтеза на память привел к предположению, что формирование памятного следа связано с активацией определенных генов при деспирализации соответствующих участков цепи ДНК. Это происходит под влиянием ионных сдвигов, отражающих возбуждение нервной клетки при поступлении в нее сигналов.
На основании того, что снятие гистоновой блокады дерепрессирует гены, обусловливающие модификацию свойств клонов клеток была предложена нейроселективная теория долговременной памяти (В.В. Дергачев, 1967). Согласно этой теории при запоминании происходит селективная дерепрессия генов, реализующая определенную комбинацию из закодированных в геноме клонов нейроглиальных комплексов. В этих комплексах более лабильная функциональная ДНК программирует такое направление синтеза РНК и белков, которое определяет специфические свойства избирательной возбудимости нервных клеток. О значении ДНК в формировании долгосрочной памяти свидетельствуют опыты, показывающие нарушения памяти при действии ингибиторов синтеза рибонуклеиновых кислот даже в то время, когда синтез белков продолжался.
Электрофизиологические исследования механизмов привыкания нервных клеток к действию раздражителей привели к представлению, что памятный след отражает не столько временные, сколько пространственные характеристики сигналов. Он представляет собой не «паттерн импульсов», а «паттерн синапсов», кодирующих своим расположением и состоянием значение сигнала (Е.Н. Соколов, 1969). При этом медиаторы через рецепторы синаптических мембран вызывают активацию одних и инактивацию других оперонов молекулы ДНК. В результате формируется система обеспечения синтеза специфических белков, реализующих функцию «обученной нервной клетки (рис. 25).
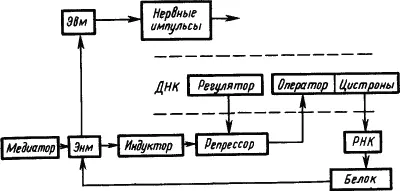
Рис. 25. Участие функциональных структур генетического аппарата молекулы ДНК в долговременной памяти (по Е.Н. Соколову):
пунктиром обозначен участок молекулы ДНК; Эвм — электровозбудимая мембрана, Энм — электроневозбудимая мембрана
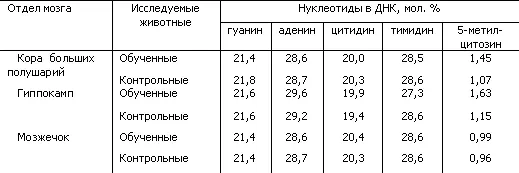
За последнее время получены новые доказательства участия ДНК в механизмах долговременной памяти. Показано, что при выработке условных рефлексов происходят своеобразные изменения структуры молекулы ДНК в нервных клетках мозга крыс. Эти изменения состояли в увеличении степени метилирования ДНК, повышающего активность определенных генов. Они были четко выражены в коре больших полушарий и гиппокампе и оказались незначительными в мозжечке. При такой «мягкой» обратимой модификации ДНК ее нуклеотидный состав оставался неизменным (табл. 5).
Таблица 5. Содержание нуклеотидов и метиловых компонентов в ДНК из разных отделов мозга у обученных и контрольных крыс (по Н.А. Тушмаловой)
Как уже было упомянуто, основные структуры нервной клетки, реализующие ее функции, состоят главным образом из белковых соединений. Поэтому многие исследователи ведут изучение белков как возможного субстрата долговременной памяти. Усиленный синтез белка при обучении дает основания для суждений о его роли в явлениях памяти. Чтобы выяснить, какие именно белки усиленно синтезируются для обеспечения процессов фиксации памятного следа, предпринималось изучение их раздельных фракций.
Исследование белков мозга голубей выявило, что при их обучении происходит возрастание фракции мембранных гликопротеидов, обозначенных 10 B и 11 A . Эти белки благодаря своим углеводным компонентам могут регулировать активность возбудимых мембран и таким образом участвовать в процессах памяти. При обучении крыс применять непредпочитаемую лапу для добывания пищи последующее дробное фракционирование мозговых белков показало особое положение встречающихся только в нервной ткани белков S-100 и 14-3-2. Конформационные изменения S-100 при связывании с ионами Ca влияют на проницаемость мембран, в частности синаптических, определяя их функциональное состояние.
Значение белков в механизмах формирования долгосрочной памяти продемонстрировано в опытах, где угнетение синтеза белка нарушало переход краткосрочной памяти в долгосрочную. Так, введение ингибитора белкового синтеза пуромицина золотым рыбкам сразу после обучения стирало его следы, но введение пуромицина через 1 ч не вызывало такого действия. Эти опыты были повторены и на других животных, показывая, что синтез белка необходим для консолидации памяти, а когда памятный след закреплен, белок уже не столь нужен.
Какие же именно белки разрушаются под действием ингибитора, препятствующего формированию долговременной памяти? Для ответа на этот вопрос исследовали разные фракции мембранных белков синаптического аппарата после нарушения долговременной памяти пуромицином. Показано, что этот ингибитор наиболее резко тормозит синтез кислых белков, к которым относится специфический для тканей мозга белок S-100. Оказалось также, что белки наиболее чувствительной к ингибитору фракции по своему аминокислотному составу принадлежат к группе внутренних белков мембраны, которые связаны с ее функциями как холинорецепторы. Это соответствует мнению, что носителями долгосрочной памяти являются холинэргические синапсы. Однако имеются указания на участие в механизмах долгосрочной памяти и адренэргической системы. Отмечено, что положительное подкрепление условных рефлексов, например пищевых, связано с функционированием преимущественно серотонинэргических, а отрицательное при оборонительных рефлексах — норадренэргических структур мозга. О возможных нейрохимических компонентах подкрепления существуют разные мнения: сюда относят и норадренэргическую систему, и дофаминэргическую. За последнее время накоплено большое количество фактов, показывающих, как участвуют нейрохимические механизмы в процессах обучения и памяти.
Другой подход к выяснению роли белков в долговременной памяти состоял в прослеживании включения в них меченых аминокислот и изменений этих белков при обучении. Так, внедрение меченного тритием лейцина в белки мозга крыс после обучения возрастало и сопровождалось увеличением числа рибосом, связанных с мембранами. При переучивании крыс способу добывания пищи на стадии бурной двигательной активности происходило даже снижение включения меченого лейцина в белки мозга. По мере усвоения нового навыка включение меченого лейцина в белки, особенно специфические, возрастает. После выработки нового навыка п<...пропуск части слова...>ния у золотых рыбок отмечен синтез трех новых фракций белков, обнаруженных с помощью меченого валина. У крыс, обученных дифференцированию зрительных сигналов для решения лабиринтной задачи, наблюдали ускоренное включение меченого тирозина в мембранные белки. Выработка условных рефлексов у голубей сопровождалась усилением синтеза гликопротеинов по показателям включения меченого валина.
Читать дальшеИнтервал:
Закладка: