Александр Коган - Основы физиологии высшей нервной деятельности

- Название:Основы физиологии высшей нервной деятельности
- Автор:
- Жанр:
- Издательство:Высшая школа
- Год:1988
- Город:М.
- ISBN:5-06-001444-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Коган - Основы физиологии высшей нервной деятельности краткое содержание
Учебник состоит из двух частей: общей физиологии высшей нервной деятельности, где изложены общие свойства условных рефлексов и нервных процессов, лежащих в их основе, и частной физиологии высшей нервной деятельности, где рассмотрены особенности условно-рефлекторного поведения основных классов животных, а также высшая нервная деятельность человека. 2-е издание (1-е — 1959 г.) значительно обновлено; особое внимание обращено на возможности использования высшей нервной деятельности для решения практических задач.
Максимальная ширина картинки в этой fb2-книге — 600 пикселей, но большинство картинок имеют размер не более 460 пикселей.
Основы физиологии высшей нервной деятельности - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
При изучении возможных причин стойкого повышения синаптической передачи было обращено внимание на медиаторы возбуждения и прежде всего на ацетилхолин. Найдено, что обучение сопровождается повышением активности фермента холинэстеразы, разрушающей ацетилхолин, а вещества, подавляющие действие холинэстеразы, вызывают нарушения памяти. В опытах с обучением крыс доставать пищу из узкой трубки определенной передней конечностью активность холинэстеразы возрастала только в контралатеральном полушарии, что подтверждает ее значение в формировании памятных следов.
Сопоставление данных о взаимосвязанной динамике ацетилхолина и холинэстеразы привело к заключению, что повышенная проводимость может возникать и устойчиво поддерживаться некоторым равновесным отношением их синтеза. Избыточная активность холинэстеразы без возрастания количества ацетилхолина не позволяет ему достичь концентрации, необходимой для деполяризации постсинаптической мембраны синапса, а увеличение количества ацетилхолина при малой активности холинэстеразы приводит к глубокой неколеблющейся деполяризации — в обоих случаях проведение импульсов через синапс оказывается блокированным. Существует предположение, что ацетилхолин может транспортироваться через синаптическую щель по транссинаптическим тяжам, которые образуются из соединения белковых мицелл, формирующихся на поверхности пре- и постсинаптической мембран.
Кроме холинэргических имеются и иные синапсы, где медиаторами служат другие вещества. С учетом всех этих сведений была составлена обобщенная схема стойкого облегчения медиаторных процессов в синапсе. На рис. 24, А показано исходное состояние синапсов. Если возбуждение пресинаптического нейрона (рис. 24, Б ) вызывает усиленное выделение медиатора, а возбуждение постсинаптического — разрушающего медиатор фермента (рис. 24, В ), то при сочетаниях условного и безусловного раздражителей происходит конвергенция импульсных потоков на синапсах (рис. 24, Г ), которая обусловливает оптимальный баланс активности медиатора и разрушающего его фермента на стабильном высоком уровне, а также активацию сети белковых тяжей, уменьшающих просвет синаптической щели (рис. 24, Д ). Показано, что повышение эффективности синапсов сопровождается конформационными изменениями мембранных белков, специфичными для синаптических мембран.
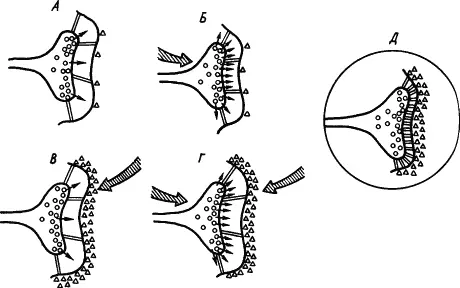
Рис. 24. Развитие процессов стойкого облегчения проводимости сигнала (объяснение см. в тексте) (по В.В. Дергачеву):
кружками обозначены везикулы медиатора, треугольниками — фермент, расцепляющий медиатор
Долгосрочная память как стойкое изменение химизма нервных клеток.Проведение сигналов от клетки к клетке, вызывающее активацию нейронных популяций, происходит в синапсах, однако функциональное состояние синапсов и свойства их деятельности в значительной мере определяются процессами, происходящими в соме клетки, в ее нейроплазме и ядре. Общеизвестна роль сомы в трофическом обеспечении передачи импульсов по путям их проведения и зависимости функционирования нейрона от его метаболизма. Это послужило основанием для поисков долгосрочного памятного следа в стойких изменениях химизма нейроплазмы.
Так как основным субстратом функциональных структур нейрона являются белковые соединения, а их различные модификации задаются матрицами РНК, на которых они синтезируются, то была выдвинута гипотеза о том, что информация, входящая в долгосрочную память, «записывается» структурой полинуклеотидной цепи молекулы (X. Хиден, 1959). По этой гипотезе разные условные потоки импульсов создают различное распределение ионных концентраций вдоль молекулы РНК и вызывают специфические для каждого сигнала перемещения нуклеотидов в их цепи. В результате каждый сигнал получает определенный отпечаток в структуре молекулы РНК. Поскольку структура молекулы РНК является матрицей для синтеза белковых соединений, специфические белки и медиаторы могут синтезироваться лишь по тому сигналу, который сформировал эту матрицу своим импульсным потоком. Все возможные перестановки и комбинации нуклеотидных элементов позволяют зафиксировать в молекулах РНК громадное количество информации. По приблизительному подсчету их информационная емкость достигает 10 15–10 20бит, что перекрывает объем человеческой памяти.
В качестве экспериментального подтверждения гипотезы о переопределении нуклеотидов в молекуле РНК как механизме формирования долгосрочной памяти могут служить результаты опытов с крысами, которые обучались добираться к пище, балансируя на проволочном мостике (табл. 4).
Таблица 4. Процентное содержание нуклеотидов в РНК ядра Дейтерса при выработке двигательного навыка у крыс (по X. Хидену, Е. Эльхази)
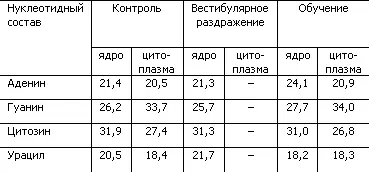
Как показывает табл. 4, у крыс, обучившихся сложным движениям, в нервных клетках ядра Дейтерса, ответственного за выполнение нового двигательного навыка, произошло изменение нуклеотидного состава РНК клеточного ядра. Увеличилась доля аденина, уменьшилась доля урацила. Простое вестибулярное раздражение не изменяло нуклеотидный состав. Исследование глиальных клеток ядра Дейтерса выявило сходные изменения РНК, что позволило сделать предположение об участии глии в кодировании поступающих сигналов путем изменения структуры нуклеотидных цепей РНК.
Однако гипотеза о нуклеотидном коде памятного следа встретилась со многими затруднениями, из которых главное — недолговечность молекул РНК. Выполняя информационные и транспортные функции, они могут сохранять свою структуру лишь в течение короткого времени, не соизмеримого с длительностью долгосрочной памяти. Эти и другие возражения ставят под сомнение РНК как носителя долгосрочного памятного следа, с чем, в сущности, согласился сам автор нуклеотидной гипотезы (X. Хиден, 1969).
В связи с этим внимание исследователей обратилось к более долговечным химическим компонентам структурно-функциональной организации нервной клетки. Памятный след стали искать в генетическом аппарате, который управляет деятельностью нервной клетки и определяет свойства на протяжении всей ее жизни. Наиболее стабильной структурой генетического аппарата являются молекулы ДНК, в которых кодируется наследственная информация. Логично было думать, что и информация, приобретаемая в течение жизни фиксируется этими структурами, специально приспособленными к хранению информационных программ. Поэтому были предприняты исследования ДНК как носителя долговременной памяти.
Читать дальшеИнтервал:
Закладка: