Феликс Филатов - КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле.

- Название:КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле.
- Автор:
- Жанр:
- Издательство:ОАО ордена Знак почета Смоленская областная типография им. В.И.Смирнова
- Год:2011
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Феликс Филатов - КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле. краткое содержание
КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле. - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
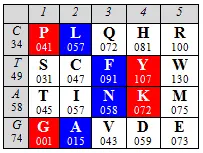
сводит число возможных версий трехмерной модели кода к единственной:
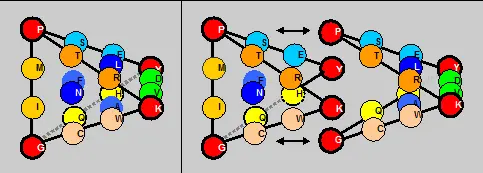
Правда, равновесную по граням модель можно также построить, заменив ребра QHи VDна QVи Н Dи сохранив, таким образом, симметричный рисунок реберных мономеров в составе матрицы, однако, эта версия потребует сделать формулировку принципа сборки тетраэдра более свободной, поскольку наш тетраэдр характеризуется также полной симметрией по граням аминокислот двух арс-классов:на каждую его грань приходится равное (по 5) число мономеров-аминокислот каждого класса.
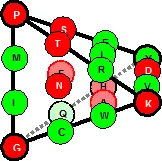
Других столь же простых условий сборки тетраэдра с нуклонным равенством граней не существует. Также (естественно) не удается сформировать подобный тетраэдр, используя значения порядковых номеров этих мономеров в качестве их альтернативных параметров. Количественная симметрия имеет место только в отношении номеров инвариантных мономеров сплошной последовательности аминокислот (независимой от арс-класса): суммы номеров мономеров вершин полученного тетраэдра и центров его граней равны (и в случае нумерации по нарастанию нуклонной массы составляют замечательное — в контексте этой и предыдущей глав — десятичное число 37).
Инвариантные мономеры и сами по себе обладают целой серией собственных симметрий по первым, вторым и третьим основаниям своих кодонов, что является следствием их положения в составе матрицы кода. Читатель может самостоятельно организовать и проанализировать таблицы, необходимые для демонстрации этих симметрий.
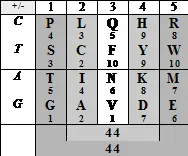
В формате позиционных номеров аминокислот, принадлежащих к тому или другому арс-классу, отметим, что значения колоночных и построчных суммацийматрицы генетического кода имеют весьма замечательный вид: линейное нарастание сумм порядковых номеров центральных колонок, выраженное двух- или трех-значнымиинфрмационными символами, в комбинации со сдвиговой для трехзначных чисел (или зеркальной для двузначных) симметрией цифр в крайних колонках, а также в строках, соответствующих первым комплементарным основаниям кодонов:
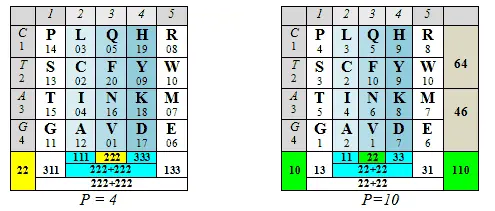
Читателю предлагается обратить внимание на следующее:
— цифровые гомодублеты, как носители информации о симметрии, имеют здесь не меньшее значение, чем гомотриплеты;
— в тех случаях, когда значения линейно нарастающих сумм порядковых номеров центральных колонок в той или иной системе счисления не подчеркнуто гомодублетами 11-22-33или гомотриплетами 111-222-333,цифровые симметрии сумм крайних колонок «комплементарны», так что сумма двух крайних колонок представляет собой следующий гомодублет (44) или гомотриплет (444);
— в тех случаях, когда значения сумм колонок или строк имеют неодинаковое число разрядов в той или другой системе счисления, цифровые симметрии сумм крайних колонок невыразительны;
— то обстоятельство, что таблица справа (где арс-классы представлены как самостоятельные группы, то есть члены каждой имеют собственную, а не последовательную, нумерацию) демонстрирует симметрии не только по колонкам, но и по строкам, свидетельствует об определенной независимости классов;
— наконец, то обстоятельство, что обе таблицы демонстрируют не только числовые, но и цифровые симметрии, как будто поддерживает логику сравнения числового значения и цифрового порядка, которые — имея общее выражение — характеризуют описанный в предыдущей главе виртуальный олигопептид.
Паттерн этих матричных симметрий в значениях порядковых номеров аминокислот сходен с паттерном симметрий матрицы в значениях их нуклонных масс (см. выше):
.....................
И хватит, пожалуй. Автору не хочется больше надоедать Читателю похожими друг на друга таблицами и «интересными» числами. Пора подумать, что все они означают. Сумбур и брызги негромкого, но явственно различимого ритма Музыки Сфер ? Сто лет назад Бернард Шоу (одно время музыкальный критик) — словно предчувствуя расцвет сегодняшней глянцевой «звездятины» — заметил, что музыка — это алкоголь осужденных грешников [62]. Справедливо, конечно, если скорбеть животом в такт второй симфонии Бетховена (http://galicarnax.livejournal.com/25217.html). Но так ли уж нужны стигматы святой Терезе ? Так ли они ей желанны ?
Однако, ничего не знал Бернард Шоу о генетическом коде, а известный вопрос Уильяма Блейка, странным образом о нем (коде) напоминающем, не мог прийти поэту в голову иначе, как в ассоциации с бенгальским тигром:
What immortal hand or eye, Dare frame thy fearful symmetry?
А нам сегодня — мог, и ничего страшного ( fearful ) или рокового , судьбоносного (fateful) — достаточно чуть-чуть «буквализировать» великолепный перевод Самуила Маршака:
Кем задуман роковой Соразмерный образ твой?
Единственное исключение из общей структуры аминокислот представляет собой пролин Р. Он содержит боковую цепь с двумя связями — и на один атом водорода меньше в константной части. Однако, воображаемая передача одного нуклона из боковой цепи — константнойвозвращает последней стандартную массу: 73+1=74, в то время, как боковая цепь остается без протона: 42-1=41. Одновременно пролину возвращается «стандартная» структура аминокислоты, состоящая из константной и вариабельной частей.
Остается отметить, что пунктуационный знак «стоп», не кодирующий аминокислоты (в отличие от знака «старт», ассоциированного с метионином М), принимается в оцифровке генетического кода за ноль. Примем также во внимание и то, что
— оцифровка генетического кода сама по себе,
— разделение молекулы аминокислоты на константную и вариабельную части для наглядности такой оцифровки,
— знак «0», которым в арифметике (особенно там, где речь идет о представлении числа в той или иной системе счисления), помечают не столько «пустоту», сколько «пробел»,
— «нормирование» молекулы пролина, показанное выше,
— кодирование цистеина триплетами TGH в таблице универсального кода -
все это — действия совершенно искусственные, работа ума, артефакт, не имеющий, на первый взгляд, никаких «естественных» аналогов. Но именно на них основана оцифровка генетического кода, о которой пойдет здесь речь. Ее результаты заставляют думать, что перед нами либо следствие физических законов необычной природы, либо случайные совпадения, либо конструкция, собранная по принципам, для демонстрации которых такая оцифровка адекватна.
Читать дальшеИнтервал:
Закладка:





