Феликс Филатов - КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле.

- Название:КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле.
- Автор:
- Жанр:
- Издательство:ОАО ордена Знак почета Смоленская областная типография им. В.И.Смирнова
- Год:2011
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Феликс Филатов - КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле. краткое содержание
КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле. - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Эта игрушка представляет собой додекаэдр, «кристалл», собранный из 120 тетраэдров (Автор не вдается здесь в детали её построения, их можно найти на сайте http://www.codefun.com/Index.htm). Разумеется, ничего подобного в природе нет, зато модель Рафики хорошо иллюстрирует симметрии кодирования и даже некоторые аспекты укладки белковых молекул.
Между тем число продуктов кодирования (20) дает соблазн собрать именно тетраэдр; надо только сформулировать простой принцип сборки, желательно учитывающийи кодирующие основания, и кодируемые аминокислоты, помня об упомянутом выше ограничении Эйгена: теория может быть корректной или нет; модель имеет третью возможность — оставаясь корректной, совершенно не относиться к делу (http://en.thinkexist.com/quotation/a-theory-has-only-the-alternative-of-being-wrong/354852.html a theory has only the alternative of being wrong; а model has a third possibility — it might be right but irrelevant ). Зато у модели, как и сказано в Главе 69 , есть очевидное достоинство: представляя явление в неожиданном ракурсе, она заставляет думать.
Здесь мы, однако, рассказываем не просто о геометрической симметрии генетического кода (о ней уже шла речь в Главе А ), но о моделях, симметрия которых базируется на оцифровке генетического кода, реализуемой по тому или иному принципу. Более того, этот подход привлекает нас, в первую очередь, тогда, когда такой оцифровке подвергаются оба компонента кода, а не только продукты кодирования. В конце предыдущей главы ( Глава Б 1 ) мы описали «виртуальный олигопептид», который демонстрировал равновесие совокупных нуклонных масс стандартных и вариабельных частей кодируемых продуктов. Мы обнаружили, что этот «олигопептид» имеет любопытные арифметические свойства в отношении составляющих его кодирующих оснований, которые, неожиданно подчеркивают акцентируемый Щербаком децимализм генетического кода. Параметр, выявляющий обнаруженные свойства, представляет собой простой номер каждого из четырех азотистых оснований в их упорядоченном по изменению молекулярной массы ряду. В данной главе мы попытаемся проанализировать организацию генетического кода, используя оба указанных параметра (нуклонные числа и порядковые номера) обоих компонентов кода. Если эта попытка окажется удачной, и мы найдем, что одна и та же организация кода (модель) характеризуется арифметическими симметриями по каждому из этих параметров, тогда легкомысленная готовность Автора сравнивать десятичное число 3412 и цифровой ряд 3412 по чисто внешнему сходству, может показаться Читателю не такой уж смешной.
Вернемся к матрице генетического кода , «аналоговая» версия которой описана в Главе А . Ее оцифровка в параметрах нуклонных масс («сжатая» версия — без пятой, @ -строки) реализуется упорядоченными по массе последовательностями первых кодонных оснований (по вертикали) и соответствующих им продуктов (по горизонтали); слева — аминокислоты в "нейтральной"версии ( 0), справа — в заряженной ( +/-). Под символом каждой аминокислоты — ее нуклонная масса (нуклонная масса боковой цепи ее молекулы).
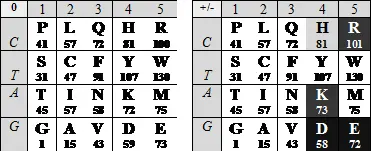
Организующая матрицу последовательность первых триплетных букв — CTAG — демонстрирует не только симметрию по комплементарности С ≡ G , A = T (черточки между основаниями символизируют число водородных связей, которые их объединяют), но и совпадающую с ней количественную симметрию цифрового ряда 1234 : 1+4=2+3. Комплементарность оснований позволяет собрать и другой ряд — AGCT , в котором упорядоченность по массе комплементарных пар имеет общее направление. Этому ряду и соответствует цифровая последовательность 3412 , описанная в предыдущей главе.
Теперь, чтобы объединить в общем представлении и аминокислоты, и азотистые основания, надо описать те и другие в общих терминах. В нашем случае это — либо нуклонная масса вариабельных частей молекулы, либо простое перечисление элементов, упорядоченных по массе. Выбор вариабельной части молекул аминокислот очевиден — это их боковая цепь. Вариабельная часть молекулы азотистого основания не представляет собой столь ясно выделяемую структуру. В то же время стандартным блоком, общим для всех оснований, является вполне выраженная структура — гексацикл (шестичленное кольцо из четырех атомов углерода, 2-4-5-6, и двух — азота 1-3):
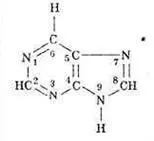
Мы подошли к выбору нуклонного параметра азотистого основания совершенно формально: все атомы вне упомянутого гексацикла и составляют нуклонное число этого основания. Таким образом, цитидину С соответствует (в полинуклеотидной цепи) нуклонная масса 34, тимину Т — 49, аденину А — 58, а гуанину G — 74. Если сопоставить комплементарные пары GC и АТ четверки нуклеотидов и их нуклонные массы, объединенные водородными связями (то есть общими для обоих членов пары протонами), получим равенство: 34+74-3 = 49+58-2 = 105 = 11 × 111 4. Стоит отметить, что все эти рассуждения относятся только к ДНК, потому что урацил U , заменяющий в четверке оснований РНК тимидин Т , имеет нуклонную массу 35, нарушающую описанные равновесия. Смысл этого обстоятельства должен отражать различия в физико-химии РНК и ДНК: возможно, дело в том, что предпочтительная структура ДНК линейна, а ее двойная спираль, уравновешенная также в описанных терминах, стабилизирует предпочтительную (линейную) запись генетической информации. РНК — в отличие от ДНК может приобретать более разнообразную конформацию, включая такую, которая позволяет ей обладать некоторыми свойствами полипептида — например, энзиматическими. В отличие от описанного выше МПП, молекула которого обладает собственным равновесием — за счет равновесия константных и вариабельных частей аминокислот, молекула ДНК приобретает равновесие только за счет объединения в спираль двух комплементарных полимеров. Само же по себе молекулярное равновесие — это характерное свойство — если не естественных биополимеров самих по себе, — то, во всяком случае, организации их генетического кодирования.
Соответствующая (реципрокная, симметричная) операция с аминокислотами, которые уже охарактеризованы в терминах нуклонных масс, — это придание им порядковых номеров в последовательностях, упорядоченных по молекулярной массе. Мы используем два варианта такой нумерации: сплошное (прямое или обратное) перечисление аминокислот в «нейтральной» версии (а) и общее или раздельное их перечисление (также в обоих направлениях) в составе арс-классов — параллельное и анти-параллельное (б). Другие варианты Читатель может рассчитать самостоятельно; мы только предупредим его, что результаты в принципе будут однотипными.
Читать дальшеИнтервал:
Закладка:





