Стивен Роуз - Устройство памяти. От молекул к сознанию

- Название:Устройство памяти. От молекул к сознанию
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1995
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Стивен Роуз - Устройство памяти. От молекул к сознанию краткое содержание
Для нейрофизиологов и всех интересующихся проблемами биологии и психологии.
Устройство памяти. От молекул к сознанию - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
В главе 7 уже говорилось, почему исследования на аплизии приобрели стратегическое значение для нейробиологии некоторых важных форм памяти. У этого животного очень несложный поведенческий репертуар, включающий различные виды научения, а их центральную нервную систему сравнительно нетрудно картировать, так как она состоит не более чем из 20 000 нейронов, сгруппированных в ряд обособленных ганглиев, и среди этих нейронов есть очень крупные клетки, легко распознаваемые индивидуально у всех особей. Ключевым моментом в подходе Кэндела было изучение простейших поведенческих реакций у интактного животного. Речь идет об элементарном рефлексе — втягивании жабры и сифона — с явлениями привыкания и сенситизации. Эту форму поведения или ее нервный механизм можно «отделить» от других поведенческих реакций путем последовательного уменьшения группы изучаемых в эксперименте нейронов. Предельным случаем будет наблюдение над двумя определенными нейронами, которые можно отпрепарировать, а затем вызвать образование синаптической связи в лабораторной чашке. По мнению Кэндела, взаимодействие таких нейронов и их реакции на добавляемые извне нейромедиаторы можно считать моделью памяти для рефлекса в ее крайне редуцированной форме. Каковы же доводы в пользу этого?
Рассмотрим сначала сам рефлекс. Аплизия дышит с помощью жабры, которая размещается в полости на верхней (дорсальной) стороне тела. В задней части этой полости имеется вырост в виде трубки — сифон. Если прикоснуться к области вокруг жабры или сифона, они втягиваются в полость: это простой защитный рефлекс. Нервный механизм такого рефлекса включает небольшое число (около 50) сенсорных нейронов, реагирующих на тактильное раздражение упомянутой области. Эти сенсорные нейроны контактируют примерно с 20 моторными нейронами как непосредственно, так и через промежуточные нейроны (интернейроны). Моторные нейроны расположены в абдоминальном (брюшном) ганглии и в свою очередь образуют синапсы на мышечных волокнах, втягивающих сифон и жабру. Эта относительно простая цепь схематически показана на рис. 9.2. Однократное раздражение поверхности тела около жабры или сифона приводит к их втягиванию, а при многократной стимуляции наступает привыкание; реакция на повторные стимулы постепенно ослабевает вплоть до ее полного исчезновения на некоторое время. Привыкание можно снять (дегабитуация, или сенситизация) сильным раздражением другого участка тела, например «хвоста», что приводит к полному восстановлению первоначальной реакции. Такие изменения рефлекса носят кратковременный и довольно неспецифический характер, и их следует рассматривать как формы неассоциативного научения. Наряду с этим, как показал Кэндел, возможна и выработка классического условного рефлекса, когда безусловным стимулом служит резкое воздействие на хвостовую область, а условным — слабое тактильное раздражение сифона. Слабая стимуляция обычно приводит и к слабо выраженному втягиванию сифона, тогда как после закрепления условного рефлекса такое же слабое раздражение вызывает сильную реакцию. Этот эффект сохраняется довольно долго, и поскольку здесь существует специфическая связь между стимулом и реакцией, его можно считать формой истинного ассоциативного научения.
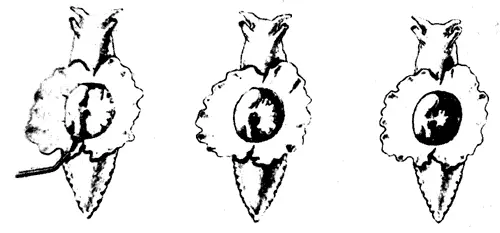
Рис. 9.2а. Рефлекс втягивания жабры и сифона у аплизии. Показано интактное животное. На сифон через трубку направлена струя воды (слева), это приводит к втягиванию сифона и фабры (в середине и справа).
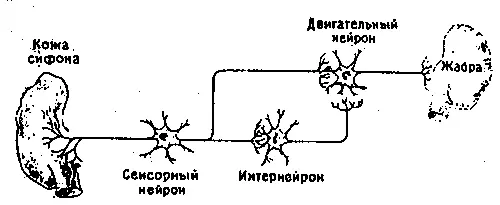
Рис. 9.2b. Схема нервных связей в «редуцированном» препарате.
Кэндел и его сотрудники занялись поисками нервных путей, участвующих в реакции втягивания жабры и сифона. Они использовали простые классические методы нейрофизиологии, а дальнейшая стратегия состояла в последовательном уменьшении числа нейронов. Для более точного контроля и количественной оценки реакции моллюсков обездвиживали, прикрепляя к пластинке, а для стандартизации тактильных стимулов применяли струю воды, направляемую через остроконечную трубку. Сокращения жабры можно было также прямо регистрировать с помощью фотоэлемента. Выявив нервную сеть, которая обеспечивала рефлекторную реакцию, исследователи смогли перейти ко второму вопросу: какая часть этой сети ответственна за привыкание? Не происходят ли при этом изменения в каких-то определенных клетках или синапсах? Этот вопрос, очевидно, имеет отношение к описанному выше первому критерию. Но, поскольку группа Кэндела состояла из нейрофизиологов, а не биохимиков, исследователи начали с изучения электрических свойств клеток, что соответствует моему шестому критерию.
Однако для получения ответа требовалось еще одно упрощение изучаемой системы, в результате которого живые аплизии превращались в неактивные, удобные для манипуляций «препараты». Животное можно вскрыть, обнажив абдоминальный ганглий, или даже полностью изолировать этот ганглий вместе с нервами, участком кожи и жаброй. Проведя такую операцию, Кэндел мог считать, что исключил все посторонние источники сигналов — другие периферические нервы, нейро-модуляторы, циркулирующие в крови и т. п. При этом он получил возможность выявлять крупные клеточные тела моторных нейронов и, как говорилось в главе 7, повторно идентифицировать «те же самые» клетки при смене одного животного другим (рис. 9.2). В результате операции живой моллюск становился чем-то вроде компьютерной схемы, и ученые могли исследовать ее свойства как электротехники, получившие новое оборудование и пытающиеся понять принципы его работы. В такой системе осязательный стимул для поведенческих реакций можно заменить его нейрофизиологическим аналогом, т. е. прямым электрическим раздражением нервов, идущих к сенсорным нейронам. Точно так же и мышечную реакцию — втягивание жабры и сифона — можно вызывать прямой стимуляцией нервов, отходящих от моторных нейронов к жаберной мускулатуре.
На следующем этапе работы изолированные и «упрощенные» препараты использовались для того, чтобы выяснить локализацию механизмов привыкания, т. е. найти те звенья нервной цепи, которые начинают слабее отвечать на повторное раздражение. К середине семидесятых годов стало ясно, что ни подходящие к сенсорным нейронам, ни отходящие от двигательных нейронов нервы не обладают свойствами, позволяющими объяснить привыкание, так как их электрические ответы не уменьшаются. Следовательно, клетки, ответственные за привыкание, должны занимать промежуточное положение в сенсорно-моторной цепи абдоминального ганглия. И действительно, регистрация электрических сигналов от моторных нейронов в этом ганглии в период привыкания выявила постепенное уменьшение их частоты. Из этого был сделан вывод, что «место» привыкания лежит между сенсорными и моторными нейронами [6].
Читать дальшеИнтервал:
Закладка:




![Станислав Сергеев - Памяти не предав: Памяти не предав. И снова война. Время войны [сборник litres]](/books/1059493/stanislav-sergeev-pamyati-ne-predav-pamyati-ne-pred.webp)