Стивен Роуз - Устройство памяти. От молекул к сознанию

- Название:Устройство памяти. От молекул к сознанию
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1995
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Стивен Роуз - Устройство памяти. От молекул к сознанию краткое содержание
Для нейрофизиологов и всех интересующихся проблемами биологии и психологии.
Устройство памяти. От молекул к сознанию - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
ДВП легко вызывать и изучать классическими методами нейрофизиологии, поэтому вряд ли стоит удивляться ее популярности в качестве потенциальной модели памяти. В ближайшие годы после первых наблюдений Блисс в Лондоне, Андерсен в Осло и все большее число исследователей в других лабораториях стали в мельчайших подробностях изучать физиологию ДВП. Было показано, что этот феномен выявляется не только у наркотизированных и ненаркотизированных кроликов, крыс и других лабораторных животных, но и в препаратах in vitro.
Гиппокамп легко извлечь из мозга вместе с его входными нервными путями, в том числе и перфорантным путем. Благодаря трехмерной структуре гиппокампа можно получать тонкие срезы с интактными путями к клеткам того же среза, как показано на рис. 9.4. Такие срезы культивировали in vitro и изучали их электрические свойства. Так забытая с пятидесятых годов методика Мак-Илвейна снова вошла в моду у нейробиологов в конце семидесятых годов. Надлежащая стимуляция нервных путей в таких срезах тоже приводила к ДВП, которая сохранялась до отмирания среза.
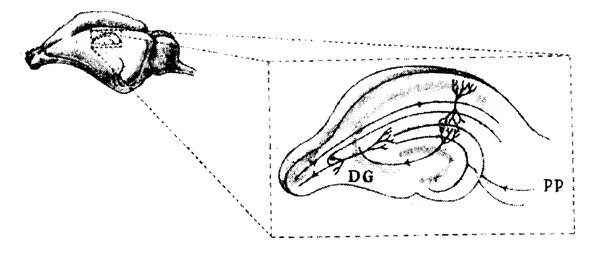
Рис 9.4. Гиппокамп крысы. На увеличенном изображении представлен срез гиппокампа с зубчатой извилиной (DG) и перфорантным нервным путем (РР) Другие области, где имеет место ДВП, включают СА1 и САЗ
Характеристики ДВП примерно одинаковы и в срезах, и в интактном мозгу. Во-первых, в обоих случаях реакция специфична для определенных путей. Иными словами, она развивается только в тех клетках, к которым подводится «условный раздражитель», а не распространяется от клетки к клетке, т. е. это результат функционирования сети специфических связей, а не волна диффузной активности. Такую специфичность можно изящно продемонстрировать благодаря наличию нескольких различных подводящих путей к отдельным участкам даже в одном срезе гиппокампа. Во-вторых, для индукции ДВП требуется повторное воздействие достаточно частых импульсов: то же число импульсов, поступающих с меньшей частотой, не вызывает эффекта. Таким образом, существует частотный порог индукции ДВП. При частоте импульсов выше пороговой ДВП может развиваться либо постепенно, либо по принципу «все или ничего» в зависимости от ритма, интенсивности и частоты условных стимулов. Развитие ДВП происходит по меньшей мере в две (возможно, в три) фазы, с коротким периодом инициации и более длительной фазой устойчивой реакции, которые рассматриваются как аналоги перехода от кратковременной памяти к долговременной.
Третий и, пожалуй, самый интересный момент с точки зрения клеточной аналогии памяти состоит в следующем. В начале восьмидесятых годов было показано, что существует ассоциативная форма ДВП. При этом слабая стимуляция, недостаточная сама по себе для поддержания ДВП, может стать активирующим фактором, если сочетается с сильной стимуляцией через второй путь [16]. Оба вида стимуляции должны совпадать или ассоциироваться во времени, как условный и безусловный раздражители при ассоциативном научении. Можно даже получить такую форму ассоциативного научения, в которой смешаны поведенческие и нейрофизиологические стимулы. Крыса может научиться переходить границу между двумя отделениями ящика в ответ на сигнал, состоящий только из залпа импульсов на гиппокамп, в качестве безусловного раздражителя [17]. Все эти особенности ДВП, взятые вместе, вызывали повышенный интерес к ее изучению по меньшей мере как одной из интригующих моделей памяти.
Гиппокамп как когнитивная карта
Признание ДВП в гиппокампе единственной моделью памяти в восьмидесятых годах скорее всего было закреплено все возраставшим потоком психологических данных о роли этого отдела мозга в обучении животных. В то время как исследования на людях позволяли предполагать участие гиппокампа в переходе от кратковременной декларативной памяти к долговременной (что подтверждалось наблюдениями над обезьянами с поврежденным гиппокампом), опыты на крысах выявили еще один аспект его роли в механизмах памяти. Одним из наиболее выраженных последствий повреждения гиппокампа у этих животных была утрата способности обучаться задачам на пространственную ориентацию, например в лабиринтах. Хотя ранее этот эффект уже наблюдали при выполнении крысами более традиционных задач с лабиринтом, его особенно убедительно продемонстрировал в своем тесте Ричард Моррис, в то время работавший в Университете Сент-Андру (сейчас — в Эдинбурге). Он использовал круглый чан с высокими стенками диаметром около двух метров, наполненный теплой водой, которую замутняли, добавляя молоко. Чан находился в комнате, на стенах которой были легко узнаваемые ориентиры: на северной стене — часы, на южной — источник света, на восточной — клетка и т. д. В чане, чуть ниже уровня жидкости, имелась полка, невидимая в мутной воде. Помещенная в чан крыса начинала беспорядочно плавать, пока более или менее случайно не натыкалась на полку и не взбиралась на нее. Путь, проделанный плавающей крысой, прослеживали с помощью телекамеры, укрепленной над чаном. После нескольких тренировок крыса стала быстро направляться почти прямо к полке, находя ее по окружающим ориентирам — часам, источнику света и клетке. Такой план эксперимента позволял легко оценивать действие различных веществ, повреждений или иных манипуляций по изменению быстроты, с которой крыса находила скрытую полку. Этот тип лабиринта стал настолько популярным, что изобретатель удостоился высшей научной почести — эпонимии (когда явление, метод или инструмент называют чьим-либо именем), а сам аппарат именуется теперь «водяным лабиринтом Морриса» (рис. 9.5). В последние годы он практически вытеснил ящик Скиннера как непременный атрибут всех психологических лабораторий.
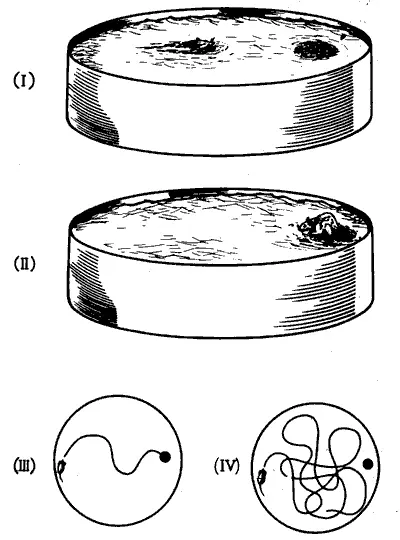
Рис. 9.5. Лабиринт Морриса. В этом тесте крысу помещают в чан с замутненной водой (I), где она учится находить находящуюся под водой площадку (II). После нескольких сеансов обучения крыса плывет сразу по направлению к площадке (III). Однако животные с поврежденным гиппокампом или после нарушения памяти химическими средствами плавает беспорядочно и лишь случайно находит площадку, как будто раньше оно не обучалось этому (IV).
Как же крыса учится находить невидимую полку? Знает ли она, скажем, расстояние, которое проплыла от точки старта, или руководствуется ориентирами, которые видит на стенах комнаты? Такие предположения легко проверить. Изменение начальной точки плавания почти не влияет на способность крысы находить полку. С другой стороны, при изменении относительного положения настенных ориентиров, например при перемещении часов с северной стены на южную, крыса теряет дорогу и плывет к той области чана, где находилась бы полка относительно часов, если бы часы оставались на прежнем месте. Следовательно, животное определяет свое местонахоядение в пространстве, используя в качестве ориентиров окружающие объекты. (Разумеется, о чем-то подобном догадается любой человек без специальной подготовки, но не так рассуждают психологи, воспитанные на заветах Скиннера). В то же время повреждение гиппокампа резко ухудшает способность крыс запоминать или вспоминать пространственные ориентиры, а тем самым и находить кратчайший путь к спасительной полке.
Читать дальшеИнтервал:
Закладка:




![Станислав Сергеев - Памяти не предав: Памяти не предав. И снова война. Время войны [сборник litres]](/books/1059493/stanislav-sergeev-pamyati-ne-predav-pamyati-ne-pred.webp)