Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Как мы помним, большая субъединица рибосомы поначалу работала в одиночку, без малой субъединицы и, следовательно, без матричной РНК. Она могла сшивать аминокислоты друг с другом, но не могла собирать из них длинные цепочки с определенной сложной последовательностью. Какие же пептидные цепи могла создавать проторибосома без матричной РНК?
Напрашивается три возможности. Во-первых, проще всего строить пептидные цепи из молекул одной аминокислоты, получая гомополимерный пептид. Во-вторых, можно использовать несколько (от двух до четырех) аминокислот, соединяя их в случайном порядке. Так образуются статистические пептиды. Третий и самый сложный вариант – когда проторибосома чередует две или больше аминокислот в постоянном порядке, производя периодический пептид.
Для гомополимерных пептидов сложно придумать какую-то функцию, которая будет востребована в мире РНК. А для периодических пептидов такие функции есть. Одну из них нам подсказывает структура рибосомных белков L2, L3, L4, L22. Эти белки состоят в основном из бета-шпилек, разделенных неструктурированными участками. Они несут положительный электрический заряд и стабилизируют укладку рибосомной РНК, компенсируя ее отрицательный заряд. Эту функцию почти так же хорошо могут выполнять периодические пептиды, полученные чередованием двухаминокислотных мотивов: (Ala-Arg) 2–4 – (Gly-Pro). Участки чередования аланина с аргинином образуют прямые части бета-шпилек, а пара глицин – пролин образует крутой поворот цепи, чтобы она могла сложиться в шпильку. Такие пептиды, которые могут встраиваться в крупные рибозимы и повышать стабильность их укладки, могли быть первым полезным продуктом пептидил-трансферазного центра. Еще более простой пример вспомогательного пептида с положительным зарядом упоминался в 10-й главе, это Tat-пептид вируса СПИДа. Этот пептид и его синтетические аналоги из повторов Arg-Gly и Arg-Gln-Gly тоже встраиваются в разные рибозимы, повышая стабильность их укладки и эффективность работы. Скорее всего, статистические пептиды такого состава тоже могут помогать работе рибозимов.
Другой вариант полезного периодического пептида можно подсмотреть в структуре активных центров различных ДНК– и РНК-полимераз. Все эти ферменты имеют в активном центре ион магния, связанный между тремя остатками аспарагиновой кислоты. Часто эти три остатка находятся на одном участке белковой цепи в составе последовательности Asp-Val-Asp-Gly-Asp. Следовательно, простое чередование аспарагиновой кислоты с валином и глицином позволяет получить вспомогательный пептид, повышающий эффективность рибозимов-РНК-полимераз.
Обойденные и вымершие аминокислоты
В предыдущей главе мы обсуждали возможные альтернативные варианты биохимии – на других химических элементах, в других растворителях, с другими генетическими молекулами вместо ДНК и РНК. Настало время обсудить происхождение набора из 20 аминокислот, составляющего белки, и оценить возможные альтернативы.
Во многих случаях природа выбрала наиболее простые аминокислоты из возможных. Аланин, например, – это самая обыкновенная аминокислота без особых примет. Глицин – единственная возможная аминокислота без боковой группы, благодаря чему по глицину белковая цепь может свободно изгибаться и вращаться. Также в стандартном наборе используются самые простые и доступные кислые аминокислоты (аспарагиновая, глутаминовая) и их амиды (аспарагин, глутамин), аминокислоты со спиртовой группой (серин, треонин), серосодержащие (цистеин), ароматические (фенилаланин, тирозин) с важной для многих ферментов имидазольной группой (гистидин) и жесткие, не допускающие вращения цепи (пролин) (Weber and Miller, 1989). Сложнее ситуация в группах положительно заряженных и неполярных аминокислот.
В стандартном наборе имеется две положительно заряженные аминокислоты – аргинин и лизин. Обе они имеют длинную боковую цепь и сложный путь биосинтеза. В аппарате Миллера и в метеоритах лизин и аргинин не встречаются, зато есть более простые аминокислоты со второй аминогруппой: диаминопропионовая, диаминобутановая и диаминопентановая (орнитин, рис. 13.9). Эти же три аминокислоты есть и в клетках. Диаминопропионовая и диаминобутановая кислоты встречаются у бактерий в составе пептидов специального назначения: антибиотиков и переносчиков железа. Орнитин является промежуточным продуктом в синтезе аргинина и есть у всех организмов – от бактерий до человека. Значит, возможно, что эти аминокислоты были доступны для включения в белки с самого начала, но по каким-то причинам вместо них были выбраны более сложные аргинин и лизин.
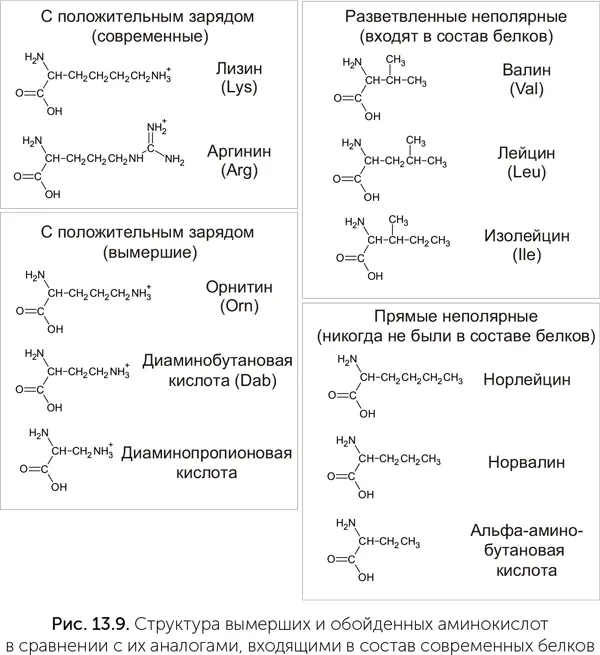
Мы можем быть уверены, что аминокислоты со второй аминогруппой входили в состав самых древних белков. Эти белки, чтобы эффективно работать в мире РНК, должны были прочно связываться с молекулами РНК, как, например, рибосомные белки. Такое связывание требует, чтобы белок нес положительный электрический заряд, т. е. имел свободные аминогруппы, которые в водной среде оказываются положительно заряжены. Рибосомные белки своим положительным зарядом компенсируют отрицательный заряд сахарофосфатного остова РНК, и только благодаря им рибосомная РНК длиной более 3000 звеньев может свернуться компактно (Hartman and Smith, 2014).
И аргинин, и лизин имеют длинные и сложные пути биосинтеза, состоящие из восьми-девяти стадий, и маловероятно, что их синтез мог проходить без участия полноценных белковых ферментов. Орнитин и диаминобутановая кислота образуются в три стадии из глутаминовой и аспарагиновой кислот, а диаминопропионовая и вовсе в один шаг из серина, т. е. они явно доступнее. Более того, для лизина у разных групп микробов существуют две разные аминоацил-тРНК-синтетазы, относящиеся к двум разным семействам. Такое разнообразие аминоацил-тРНК-синтетаз может означать, что они появились независимо, а значит, лизин вошел в генетический код уже после расхождения крупных групп микробов. Итак, у нас есть три факта:
1) положительно заряженные аминокислоты должны были быть в белках с самого начала;
2) современные аминокислоты этой группы – лизин и аргинин – имеют очень длинный и сложный биосинтез и могли возникнуть в эволюции позже других аминокислот. Для лизина это подтверждается и разнообразием аминоацил-тРНК-синтетаз;
3) существуют более простые и доступные аминокислоты с положительным зарядом, такие как орнитин, они есть в клетках, но не входят в состав белков.
Возникают подозрения, что простые положительно заряженные аминокислоты когда-то входили в состав белков, но были затем вытеснены лизином и аргинином.
Читать дальшеИнтервал:
Закладка:






