Умберто Матурана - Древо познания

- Название:Древо познания
- Автор:
- Жанр:
- Издательство:Прогресс-Традиция
- Год:2001
- Город:Москва
- ISBN:5-89826-103-6
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Умберто Матурана - Древо познания краткое содержание
Авторы книги — известные во всем мире выдающиеся чилийские нейробиологи, предложившие в начале семидесятых годов радикально новую концепцию живых систем от (простейших до человека), их происхождения и взаимосвязей. Написанная живо и интересно, с блестящим педагогическим мастерством, книга «Древо познания» делает читателя участником становления новой междисциплинарной научной парадигмы, приобщает его к новым взглядам на язык и его роль в эволюции и человеческой коммуникации. В основе этой парадигмы лежит принцип: «Живые системы — это познающие системы, а жизни, — это процесс познания».
Книга Франсиско X. Варелы и Умберто Р. Матураны "Древо познания" впервые увидевшая свет в конце 80-х годов, получила широкую международную известность в качестве интеллектуального бестселлера. Написанная первоклассными учеными, крупнейшими нейрофизиологами современности, она позволяет по-новому взглянуть на биологическую эволюцию и человеческое познание. Читатель знакомится с развитой авторами оригинальной концепцией аувтопоэза, занявшей центральное место в исследованиях современной психотерапии, стратегического менеджмента, процесса глобализации
Древо познания - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Мы подчеркиваем, что тесная агрегация клеток, происходящих от одной клетки, агрегация, которая превращается в метаклеточное единство, есть условие, которое согласуется с непрерывным аутопоэзом зтих клеток. Однако такие клеточные скопления не являют ся биологически необходимыми, поскольку многие организмы на протяжении долгой истории своего существования оставались одноклеточными. В тех же наследственных рядах где клеточная агрегация произошла, она привела к глубоким последствиям для соответствующих историй структурных преобразований. Рассмотрим эту ситуацию более подробно.
Ясно, что онтогенез метаклеточной системы должен определяться областью взаимодействий, задаваемых ею как единым целым, а не отдельными взаимодействиями образующих ее клеток. Иначе говоря, жизнь многоклеточной особи как единства протекает через функционирование ее компонент, но не определяется их свойствами. Но каждая из многоклеточных особей возникает в результате деления и обособления наследственных линий клеток, которое начинается в момент оплодотворения одной клетки, или зиготы, порожденной некоторыми органами или частями многоклеточного организма. Если новые особи не рождаются, то наследственная линия обрывается. А для того, чтобы новые особи рождались, их возникновение должно начинаться с одной клетки. Все происходит по следующей простой схеме: логика строения каждого многоклеточного организма требует, чтобы он был составной частью цикла, в котором с необходимостью существует одноклеточная стадия.
Именно на стадии одноклеточной репродуктивной фазы многоклеточного организма происходят изменения, передающиеся в поколениях. Поэтому не существует принципиальных отличий между путями становления наследственных линий у одно- и многоклеточных организмов. Иначе говоря, жизненный цикл многоклеточного организма представляет собой единство, в котором онтогенез заключается в переходе от одноклеточного к многоклеточному состоянию, но репродукция и репродуктивные изменения происходят на одноклеточной стадии.
Все известные многоклеточные организмы представляют собой искусные вариации на одну и ту же тему: клеточная организация и построение филогении. Каждая многоклеточная особь представляет собой тщательно подготовленный этап в онтогенезе наследственного ряда, тогда как его изменения продолжают происходить на клеточном уровне. В этом отношении многоклеточность не вносит ничего принципиально нового. Новизна состоит в том, что многоклеточность делает возможным возникновение множества различных классов особей, поскольку становятся возможными многочисленные наследственные линии, использующие разнообразные способы сохранения онтогенетической структурной связи в окружающей среде. Неисчерпаемое разнообразие живых существ на Земле, включая нас с Вами, обусловлено появлением многоклеточного варианта в рамках клеточных наследственных рядов.
Вместе с тем следует заметить, что половое размножение многоклеточных организмов не изменяет основную характеристику репродукции, с которой мы познакомились в предыдущей главе. Действительно, половое размножение требует, чтобы одна из клеток многоклеточного организма стала дополнительно обладать независимой операциональной динамикой (как сперматозоид) и слилась с другой клеткой другого организма того же класса, образуя зиготу, т. е. одноклеточную фазу этого же организма. Существуют также многоклеточные организмы, которые размножаются не только половым путем, но и простым делением, или исключительно делением. В зтом случае единством, изменяющимся в чреде поколений, является не клетка, а весь организм.
Последствия полового размножения наглядно проявляются в возникающей в результате него богатой структурной рекомбинации. Половое размножение, с одной стороны, делает возможным скрещивание репродуктивных линий, а с другой — резко увеличивает число структурных изменений, возникающих в каждом репродуктивном акте. Таким способом генетика и наследственность обогащаются эффектами, возникающими в результате комбинаций структурных альтернатив, присущих различным группам живых существ. Эффект возрастания изменчивости, который (как будет показано в следующей главе) делает возможным филогенетический дрейф, объясняет, почему половое размножение столь широко распространено среди живых существ: оно способствует разветвлению наследственных линий.
Темп преобразований
Превосходный способ анализа метаклеточных систем и их жизненных циклов заключается в сравнении продолжительности периодов, за которые они совершают свои жизненные циклы, в зависимости от их размеров 2. Например, на рис. 22а изображен жизненный цикл миксомицета (на рассмотрении которого мы останавливались выше). По одной оси отложено время, за которое завершается каждая стадия, а по другой — размеры, которых организм достигает к соответствующему времени. Так, на формирование плодоносящего тела длиной примерно в 1 см уходит около одного дня. Спора размером в несколько десятков миллионных долей метра формируется примерно за 1 минуту.
Bonner J. Т. Size and Cycle. — Princeton, N.Y: Princeton University Press, 1965).
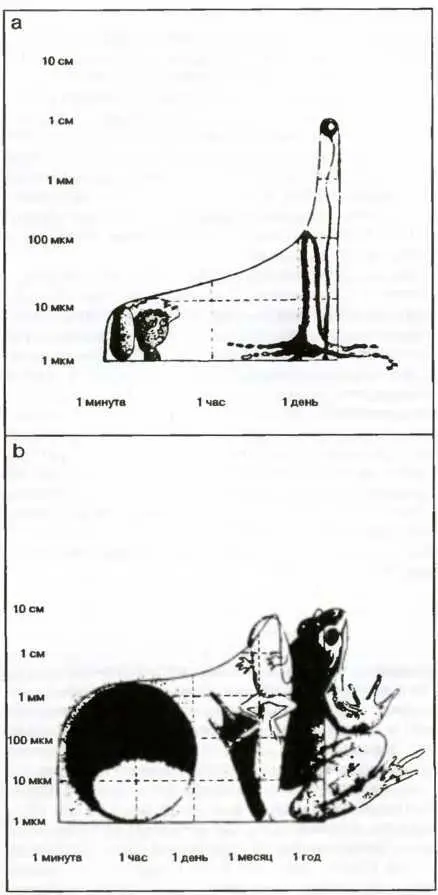
Рис. 22. Примеры зависимости между размером, достигаемым на различных стадиях жизненного цикла четырех организмов, и временем, необходимым для достижения этих стадий
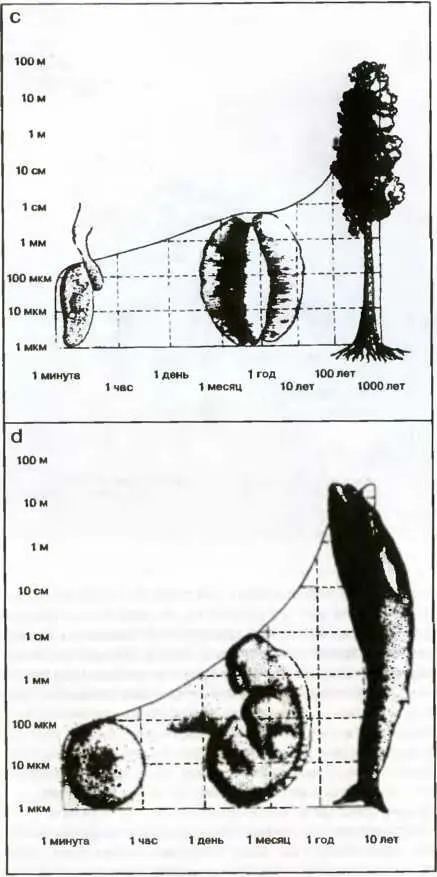
Аналогичная зависимость, на этот раз для лягушки, построена на рис. 22 Ь. Зигота, из которой затем вырастает взрослая особь, образуется примерно за минуту, в то время как взрослой особи требуется почти год, чтобы вырасти до размеров в несколько сантиметров. То же самое относится и к самому большому дереву в мире — секвойе; за время формирования, составляющее тысячу лет, она вырастает более чем на 90 метров (рис. 22 с). Это же можно сказать и относительно самого крупного в мире животного — синего кита, вырастающего за 10 лет до 40 и более метров в длину (рис. 22 d).
Независимо от размеров и внешнего вида этапы во всех перечисленных выше случаях всегда одни и те же: из начальной клетки в процессе клеточного деления и дифференциации формируется индивидуум

Рис. 23. Время репродукции одноклеточных и многоклеточных организмов второго порядка посредством сопряжения между клетками, образующимися в результате клеточных делений. Возникающая особь претерпевает онтогенез различной длительности и достигает следующей репродуктивной стадии при образовании новой зиготы. Таким образом, репродуктивный цикл является основной единицей, которая одновременно и сохраняется, и трансформируется со временем. Один из способов уяснить себе это заключается в том, чтобы построить график зависимости между временем репродукции и размерами (рис. 23). Бактерия — одноклеточное существо — размножается необычайно быстро. Следовательно, и скорость ее трансформации также высока. Одна из непременных особенностей образования единств второго порядка путем клеточной агрегации связана с тем, что для клеточного роста и дифференциации необходимо время, поэтому частота смены поколений у многоклеточных гораздо ниже.
Читать дальшеИнтервал:
Закладка:







