Владимир Живетин - Человеческий риск (системные основы управления)

- Название:Человеческий риск (системные основы управления)
- Автор:
- Жанр:
- Издательство:Изд-во Института проблем риска, ООО Информационно-издательский центр «Бон Анца»
- Год:2012
- Город:Москва
- ISBN:978-5-986640-70-9, 978-5-905883-13-2
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Владимир Живетин - Человеческий риск (системные основы управления) краткое содержание
Риски и безопасность обусловлены взаимоотношением человека с социо-природной системой при реализации потребностей человека и среды согласно возможностям человека и среды.
В монографии излагаются основные фрагменты расчета человеческих рисков на системном уровне, когда система синтезирована на структурно-функциональном уровне.
Человеческий риск (системные основы управления) - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Блок 1 – синтез РНК; экспрессия гена;
Блок 2 – энзимные, рецепторные, мембранные белки;
R 1 – расход информации из памяти, т. е. вызов из памяти нужной информации.
Физические и биохимические изменения, обеспечивающие эффективность синаптической передачи, осуществляются на основе энергетических или обменных компонентов синаптической активации в процессе обучения (жизненного цикла). Эта модель основывается на системных свойствах мозга, на топологии межнейронных связей. Главное достоинство этой модели обосновывается наличием трех главных форм памяти:
– кратковременной памяти; ее длительность измеряется секундами, она возникает в процессе переработки информации, при обучении;
– долговременной памяти; формируется из кратковременной и сохраняется от нескольких дней до конца жизни;
– промежуточной памяти, длящейся до завершения образования (консолидации) долговременной памяти.
Этим трем главным формам памяти соответствуют три главных формы регуляции синаптической связи.
В основе кратковременной памяти лежат изменения (перестройки) в пре– и постсинаптических структурах. Такие изменения вызываются либо изменениями ионного баланса, либо выделяющимися при этом нейромедиаторами. При этом происходят кратковременные изменения функций синапсов и каких-либо компонентов мембраны, которые основываются, главным образом, на конформационных перестройках мембранных блоков.
Биологическая основа промежуточной памяти характеризуется умеренными скоростями кинетики и связана с изменениями функциональных характеристик постсинаптических мембран.
Долговременная память характеризуется изменениями активности генов, т. е. количественным и качественным изменением ДНК-зависимого синтеза РНК, что влечет за собой соответствующие изменения синтеза белка. Благодаря встраиванию вновь синтезированных макромолекул в мембрану и включению их в обмен веществ длительность изменений функций синапса оказывается достаточной для развития долговременной памяти. Эта модель противостоит представлению об образовании «молекул памяти» на молекулярном уровне и основывается на системных свойствах мозга, на топологии межнейронных связей.
Теперь кратко остановимся непосредственно на блоке «память». Схема цепей обменных процессов в нервной клетке представлена на рис. 2.17. Обменные процессы развиваются под влиянием связывания дофамина со своими рецепторами и ведут к двухфазному повышению образования гликопротеидов в блоке 1. Два ряда процессов, встречаясь, приводят к образованию гликопротеида. Этот белок, достигая клеточной мембраны, меняет ее состав и свойства. При этом в памяти человека возникают следующие модели окружающего мира:
– сознание – деятельность, результатом которой являются «реальные» модели;
– подсознание – промежуточное состояние, когда модели нечеткие, размытые;
– бессознательная деятельность, в процессе которой нет реальных моделей.
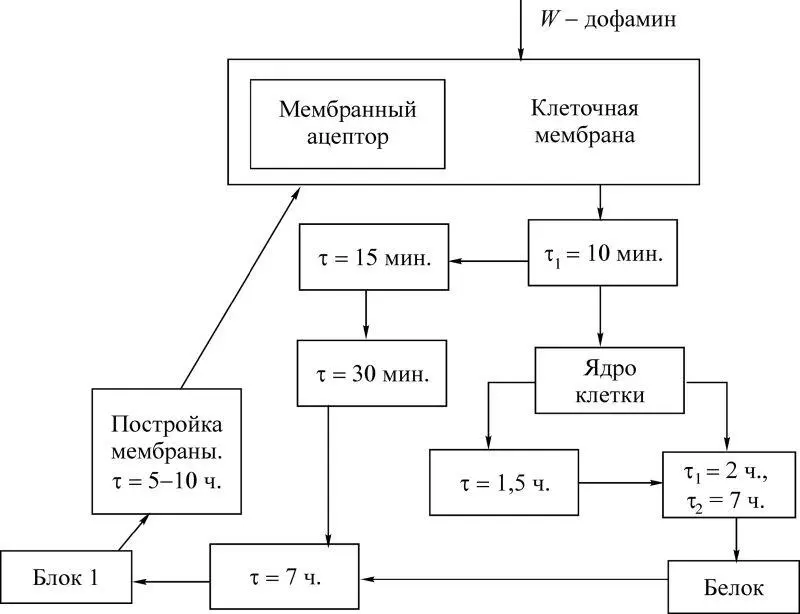
Рис. 2.17
Для анализа процесса здесь выделено главное – фактор времени, так как перестройка (топологическая) функциональных структур при развитии долговременной памяти происходит за счет образования гликопротеидов.
Кратковременная и долговременная память играют важную роль при формировании моделей окружающего мира в блоках памяти, формируя достоверные знания и знания с погрешностями, которые являются источниками риска человека.
Быстроизменяющиеся модели фактического или реального мира по-разному фиксируются у разных людей. В связи с этим можно говорить о разной величине риска, ибо более точное (т. е. более адекватное) отображение мира позволяет при малых затратах энергетики получить правильное решение и достичь цель. Пороговые величины кратковременной памяти ограничивают возможности по скорости изменения информации, поступающей от внешнего мира. Антипод кратковременной памяти – долговременная память – имеет порог по объему памяти J кр . Выход за [ x н кр , x в кр ] приводит к размытым моделям среды жизнедеятельности, что приводит к ложным решениям и неправильным поступкам и обусловливает критические состояния человека, т. е. его риск. Этот риск может быть связан как с потерей функциональных возможностей организма человека, так и с противодействием среды жизнедеятельности.
Таким образом, для регулирования величины риска человек обязан изучить области, среду, затем оценить свои возможности и необходимость привлечения дополнительных средств, прежде чем принимать решения и исполнять их. При этом клетки не являются носителями информации. Они лишь инструменты возникновения новых топологических отношений в нервной сети и, следовательно, новых пространственно-временных носителей информационных процессов.
Объем памяти у некоторых людей огромен, они могут запоминать страницы текста за короткий промежуток времени, другие – нет. Это означает, что параметры информационных блоков у разных людей различны. Под параметрами будем понимать объем памяти, быстродействие, аналитические возможности по ее обработке. Таким образом, память – это система, включающая ряд подсистем, которая имеет пороги по объему внешней информации, скорости ее поступления. Однако, как и во всякой биосистеме, в ней возможны процессы самоорганизации, перестройки и адаптации в процессе жизнедеятельности человека.
2.2.2. Нейронные сети головного мозга человека. Самоорганизация
Исследования показывают, что нервные эмбриональные центры, состоящие всего из нескольких десятков клеток культуры тканей, обнаруживают тенденцию к самоорганизации. Изолированные группы нервных клеток показывают следы своеобразной нервной активности. Эти «шумовые» «спонтанные» возбужденные состояния не ограничиваются тем нейроном, в котором они возникли, а распространяются дальше и на другие аналитические, связанные с ним нейроны.
Группы нервных клеток, изолированные от естественных контактов в нервной системе, способны организовываться в единую систему. Каждая такая группа в зависимости от количества, вида и многообразия внутренних элементов развивает разные, но определенные формы активности и поддерживает их в течение длительного времени. Важно то, что такие нейронные группы, функционирующие по принципу самоорганизации, не нуждаются в афферентном возбуждении. Соединяя эти группы (каким-либо образом) с сенсорными нейронами, легко преобразовать их в искусственную систему рефлексов, однако для возникновения процессов самоорганизации в этом нет необходимости. Так, например, изолированные центры, содержащие сегментный двигательный аппарат спинного мозга [1], способны при помощи механизма самоорганизации генерировать выходной моторный импульс, необходимый для шагающего движения конечности. Отметим, что для образования выходного сигнала, генерирующего подобное движение, достаточно даже небольшой части одного из трех-четырех сегментов спинного мозга, иннервирующих конечности. Таким образом, не существует отдельного «командного» нейрона, и не имеет смысла утверждать, что вся нейронная сеть сегментов, приводящих в движение конечность, ответственна за генерацию выходного сигнала.
Читать дальшеИнтервал:
Закладка:









