Елена Николаева - Психофизиология. Психологическая физиология с основами физиологической психологии. Учебник

- Название:Психофизиология. Психологическая физиология с основами физиологической психологии. Учебник
- Автор:
- Жанр:
- Издательство:Array Литагент «Когито-Центр»
- Год:2008
- Город:Москва
- ISBN:978-5-9292-0179-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Елена Николаева - Психофизиология. Психологическая физиология с основами физиологической психологии. Учебник краткое содержание
Для студентов высших учебных заведений, обучающихся по психологическим и биологическим специальностям. Может использоваться в учебном процессе по педагогическим и медицинским направлениям и специальностям. Представляет интерес не только для ученых и специалистов, но и для широкого круга читателей.
3-е издание, переработанное и дополненное.
Психофизиология. Психологическая физиология с основами физиологической психологии. Учебник - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:

Рис. 1.14. На высоте активности нейрона, когда потребности в ионах натрия больше, чем это обеспечивает натрий-калиевый насос, астроцит действует как насос, перекачивая натрий из ближайшего сосуда (Kalat,1992).
Когда достигается некоторое критическое значение потенциала, называемое пороговым, на уровне аксонального холмика нейрона возникает ПД – распространяющийся по аксону потенциал. При этом положительная обратная связь на уровне мембраны нейрона приводит к регенеративным сдвигам, в результате которых знак разности потенциалов изменяется на противоположный, т. е. внутреннее содержимое клетки становится заряженным положительно по отношению к внешней среде. Приблизительно через 1 мс проницаемость мембраны для натрия падает, натрий-калиевый насос выбрасывает натрий из клетки, и трансмембранный потенциал возвращается к своему значению в состоянии покоя – 70 мВ.
После каждого такого разряда нейрон становится на некоторое время рефрактерным (неспособным к активации), т. е. натриевая проницаемость мембраны в этот период не может изменяться. Это кладет предел частоте генерации ПД – не более 200 раз в секунду. Максимальная скорость распространения нервного импульса составляет приблизительно 100 м/сек. Это более чем в миллион раз меньше скорости, с которой электрический сигнал движется по медной проволоке. Таким образом, скорость распространения ПД сравнительно низка.
Синаптическая передача информации
Уже отмечалась важная роль мембраны в передаче информации в мозге. Мембрана представляет собой барьер для прохождения нервного импульса. Именно поэтому связи между нейронами опосредуются химическими передатчиками – нейромедиаторами (mediator – посредник, англ .), выделяющимися из окончаний аксонов в области специализированных межклеточных контактов – синапсов. Синапс представляет собой мембраны двух соседних нейронов (передающего информацию и получающего ее) и пространство между ними, которое называется синаптической щелью. Синаптическая щель – это пространство шириною около 20 нм между мембранами пресинаптической (мембрана нейрона, находящегося перед синаптической щелью) и постсинаптической (мембрана клетки, находящейся после синаптической щели) клеток (рис. 1.15).
Различают аксо-соматические синапсы, сформированные мембранами аксона и тела (сомы) нейрона, аксо-дендритные, состоящие из мембраны аксона и дендрита, и аксо-аксональные, при которых аксон подходит к другому аксону. Синапс между аксоном и мышечным волокном называется нейромышечной кольцевой пластинкой.
Молекулы медиатора находятся в везикулах – особых пузырьках, расположенных в аксональной терминали (окончании аксона). ПД, достигая аксональной терминали, становится сигналом открытия кальциевых каналов, которые вызывают синхронный эндоцитоз – координированное выделение медиатора из везикул и поступление их в синаптическую щель. Медиатор связывается с рецептором, находящимся на постсинаптической мембране, который инициирует в постсинаптической клетке те или иные изменения в зависимости от вида рецептора. Медиатор, взаимодействуя с рецептором, может способствовать открытию ионных каналов (натрий-калиевых или кальциевых) или через аденилатциклазный механизм активировать внутриклеточного посредника – цАМФ (циклический аденозинмонофосфат) и цГМФ (циклический гуанозинмонофосфат). При открытии натрий-калиевого канала натрий поступает внутрь клетки, что приводит к деполяризации участка мембраны постсинаптического нейрона. Каждый синапс делает лишь незначительный вклад в этот процесс. Однако каждый нейрон непрерывно интегрирует до 1000 синаптических входов, которые суммируются нелинейно (рис. 1.16) и при достижении порогового потенциала вызывают ПД, т. е. распространяющийся вдоль аксона потенциал.
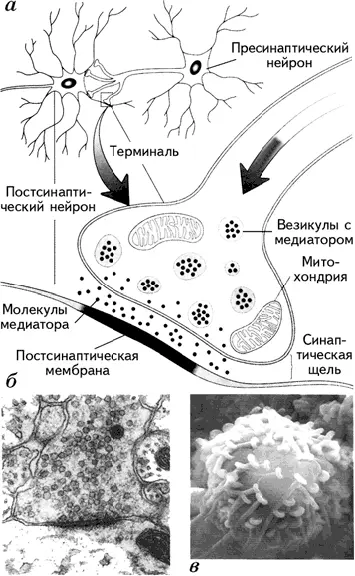
Рис. 1.15. Структура синапса (а); фотография синапса, увеличенного под электронным микроскопом в тысячи раз (выполненная D. D. Kinkel) (б); фотография аксональных терминалей на теле аксона (в) (Kalat, 1992).

Рис. 1.16. Временная и пространственная суммация импульсов (Kalat, 1992).
Синаптическая задержка – время между началом пресинаптической деполяризации и постсинаптической реакцией – составляет 0,5 мс. Всего лишь 1/10 часть этого времени обусловлена диффузией (проникновением) медиатора через синаптическую щель к постсинаптической мембране. Большая же часть времени тратится на открывание Са 2+-каналов, через которые кальций проникает в аксонную терминаль и способствует высвобождению медиатора из везикул. Кальций в аксонной терминали имеется в крайне малых количествах. После того как он окажет свое действие, он удаляется, либо связываясь со специальным белком – кальмодулином , либо проникая в эндоплазматический ретикулум. Освобожденный медиатор может действовать как на ауторецепторы (расположенные на пресинаптической мембране), так и на постсинаптические рецепторы.
Согласно принципу английского ученого Г. Дейла, постулирующего метаболическое единство нейрона, во всех окончаниях нейрона выделяется один и тот же медиатор. В настоящее время доказано, что этот принцип касается только пресинаптического единства нейрона. Эффекты, которые вызываются данным медиатором, могут быть различны и зависят от клеток-мишеней (в данном случае постсинаптических нейронов). Знак синаптического действия – повышение постсинаптического потенциала или его падение – определяется не медиатором, а свойствами рецепторов на постсинаптической клетке.
Постсинаптические рецепторы одного пресинаптического нейрона могут фармакологически различаться и контролировать разные ионные каналы. Одна постсинаптическая клетка может иметь более одного типа рецепторов для данного медиатора, и каждый из этих рецепторов контролирует отличный от других механизм ионной проводимости.
Кроме нейронов, суммирующих и передающих информацию к другим клеткам, описаны так называемые пейсмекерные нейроны , способные самостоятельно генерировать электрические импульсы (Alving, 1968). Активность таких нейронов характеризуется синусоидальными колебаниями частотой 0,1–10 Гц и амплитудой 5–10 мВ. Эти нейроны при отсутствии любого внешнего воздействия обеспечивают периодическую генерацию ПД и передачу возбуждения другим нейронам.
Читать дальшеИнтервал:
Закладка:









