Владимир Виноградов - Стресс и патология

- Название:Стресс и патология
- Автор:
- Жанр:
- Издательство:Array Литагент «Белорусская наука»
- Год:2007
- Город:Минск
- ISBN:978-985-08-0829-5
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Владимир Виноградов - Стресс и патология краткое содержание
Книга рассчитана на медиков и широкий круг специалистов, интересующихся проблемой коррекции кардио– и иммунопатологии стресса.
Стресс и патология - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
[70]. Явными проявлениями низкоэнергетического сдвига в принятых условиях являются: ослабление энергетической регуляции дыхания (рост дыхания в состоянии покоя (V 0) и снижение ДК л), рост дыхания в состоянии V 2и признаки повреждения митохондрий: относительное снижение окисления сукцината в активном состоянии (V 3), в фазе истощения иммобилизационного стресса (72 ч), которое in vitro не устраняется глютаматом, и снижение ДК чбольше, чем ДК л, во все сроки опыта.
Иммобилизационный стресс на фоне тиамина не сопровождается признаками повреждения митохондрий сердца: скорость окисления сукцината в терминальной фазе стресса (72 ч) продолжает нарастать без резкого ухудшения энергетической регуляции дыхания (имеет место относительный рост ДК л, а также одинаковое увеличение ДК ли ДК ч), которое наблюдается у стрессировавшихся крыс. Скорее всего, в принятых условиях тиамин действует как антистрессор: витаминзависимое снижение амплитуды стероидогенной реакции (рис. I-1) автоматически ограничивает степень активации сукцинатдегидрогеназы стрессорными гормонами [39]. Поскольку добавка ЩУК-устраняющего субстрата (глютамата) in vitro повышает скорость окисления ЯК митохондриями, выделенными из сердца животных, получавших тиамин во все фазы стресса, здесь возможно и другое объяснение.
Таблица I-1.
Влияние тиамина (Т) на окислительную и фосфорилирующую функцию сердца крыс в динамике ИС
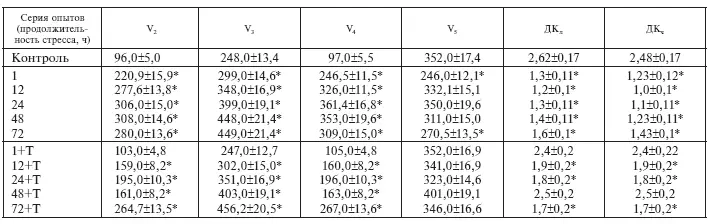
* Достоверные изменения – p < 0,05.
Согласно Г. Селье, существует 2 типа адаптационных механизмов – кататоксические, ответственные за активное сопротивление раздражителю, и синтоксические, обеспечивающие пассивную устойчивость и сосуществование с патогенным воздействием [312]. Примером перехода от кататоксических реакций к синтоксическим является тиаминзависимое ограничение окисления сукцината митохондриями сердца крыс (V 3) в динамике истощающего иммобилизационного стресса (табл. I-1). Фактически это ограничение представляет собой синтоксическую реакцию, которая и обеспечивает сосуществование с раздражителем, повышая пассивную устойчивость за счет снижения активных реакций.
Для иммобилизационного стресса показано, что на уровне митохондрий ЩУК-обусловленное ограничение дыхания выполняет функцию синтоксической реакции, предупреждая кататоксическую гиперактивацию окисления сукцината [70]. Известно, что увеличение доли ЯК в общем окислении обеспечивает повышенные энергетические запросы при активности. Однако эта компенсаторная реакция подобно адаптационным реакциям на уровне организма может становиться чрезмерной и повреждающей. В условиях повышенного содержания жирных кислот и ионов Са 2+, характерных для стресса, повышается проницаемость мембран митохондрий и возрастает интенсивность окисления субстратов. При этом активируется сукцинатдегидрогеназа и гиперактивное окисление сукцината становится источником дальнейшего повреждения мембраны [69]. На энергизованных митохондриях печени сукцинат обычно оказывает стабилизирующее действие [15], а повреждение органелл при его окислении наблюдается на интенсивно метаболизирующих низкоэнергизованных объектах, таких, как митохондрии сердца голубя и митохондрии патологического сердца человека [148]. Считается, что повреждающее действие ЯК, проявляющееся снижением или потерей дыхательного контроля, обусловлено накоплением протонов и гиперактивным транспортом ионов кальция в митохондрии.
Кальцификация тканей характерна для стресса, и она базируется на гиперактивном окислении сукцината. Интенсификация окисления этого субстрата и транспорта Са 2+в митохондрии наблюдается в ткани сердца человека при тяжелых формах сердечной недостаточности [148]. Залповый импорт кальция в кардиомиоциты после ишемизации сердца ответствен за повреждение миокарда [98]. В таких условиях блокирование дыхательной цепи цианидом предотвращает чрезмерное поступление внешнего кальция в кардиомиоциты и их повреждение [69]. Поскольку тотальное ингибирование дыхательной цепи не может быть использовано в целостном организме для in vivo профилактики повреждения митохондрий сердца крыс, при длительной иммобилизации животных некоторые авторы применяли введение антагониста катехоламинов – серотонина, который снижает дыхание тканевых препаратов [81] предположительно за счет индукции ЩУК-механизма ограничения окисления сукцината и тем самым обеспечивает защиту органелл от стресса [69]. Это допущение согласуется с повышением уровня серотонина в организме при таких типичных проявлениях хронического стресса, как язвенная болезнь желудка и 12-перстной кишки, раздражении электротоком, возбуждении, хирургических операциях и других стрессобусловленных патологических состояниях [81]. Не исключено, что инициация ЩУК-ограничения окисления сукцината митохондриями сердца при стрессе является характерным моментом действия любого антистрессора, в том числе и тиамина.
В присутствии динитрофенола, вызывающего полное разобщение дыхания и фосфорилирования, потребление кислорода митохондриями животных в терминальной фазе истощающего стресса (72 ч опыта) снижено в 1,75 раза (табл. I-1). Это означает, что изменения, выявляемые в митохондриях, изолированных из сердца животных при длительной иммобилизации, выражаются не только в нарушении сопряжения окисления с фосфорилированием, но и в нарушении самого окисления, т. е. транспорта электронов в дыхательной цепи. В соответствии с современными представлениями такие нарушения могут быть обусловлены повреждением липидного бислоя митохондриальных мембран продуктами перекисного окисления липидов (ПОЛ), а также чрезмерной активацией фосфолипаз сердечной мышцы и детергентного действия избытка жирных кислот, возникающего при стрессе в результате повышения секреции катехоламинов [98]. Как видно из табл. І-2, с увеличением экспозиции иммобилизации крыс содержание в сердце продуктов ПОЛ (диеновые конъюгаты, малоновый диальдегид) неуклонно возрастает (с максимумом к 72 ч опыта), а уровень антиоксидантной защиты кардиомиоцитов (активность каталазы, супероксиддисмутазы и содержание эндогенного токоферола) пропорционально падает, т. е. имеет место прогрессирующее нарушение исходной сбалансированности между ферментными системами генерирования и детоксикации липопероксидов за счет стрессзависимого снижения антиоксидантного статуса организма [138].
Предварительное введение нетоксичных доз природных и синтетических антиоксидантов (токоферол, ионол) предупреждает типичную для тяжелого ЭБС активацию ПОЛ в мышце сердца, мозге и других органах. В результате ингибирования ПОЛ не развивается повреждений мембран кардиомиоцитов, нарушения работы Санасоса, окислительного фосфорилирования в митохондриях, избыточной потери миокардом ферментов, транзиторного повреждения и последующей репарации ДНК; предотвращаются депрессия сократительной функции миокарда и постстрессорное снижение устойчивости сердца к гипоксии. Эти факты свидетельствуют о том, что активация ПОЛ действительно составляет ключевое звено патогенетической цепи стрессорной альтерации кардиомиоцитов. Блокирование ПОЛ, устраняющее возможность накопления гидроперекисей липидов в мембранных структурах кардиомиоцитов, очевидно, приводит к стабилизации их липидного бислоя и этот мембранотропный эффект составляет суть кардиопротекторного действия антиоксидантов [98]. В механизме снижения уровня ПОЛ под влиянием антиоксидантов, содержащих гидроксильные группы фенольного типа, наряду с осуществлением их скевенджерной функции «ловушек» супероксидных радикалов важную роль может играть и собственная антистрессорная активность препаратов.
Читать дальшеИнтервал:
Закладка: