БСЭ БСЭ - Большая Советская Энциклопедия (ФО)

- Название:Большая Советская Энциклопедия (ФО)
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
БСЭ БСЭ - Большая Советская Энциклопедия (ФО) краткое содержание
Большая Советская Энциклопедия (ФО) - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Д×Н 2+ А 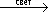 AH 2+ Д,
AH 2+ Д,
где AH 2– продукты Ф.
Структурные особенности фотосинтетического аппарата.Высокая эффективность Ф. высших зелёных растений обеспечивается совершенным фотосинтетическим аппаратом, основа которого – внутриклеточные органеллы – хлоропласты (в клетке зелёного листа их 20–100). Они окружены двуслойной мембраной. Внутренний слой её построен из уплощённых мешочков или пузырьков, называемых тилакоидами, которые часто упакованы в стопки, составляют граны, соединённые между собой одиночными межгранными тилакоидами. Тилакоиды состоят из собственно фотосинтетических мембран, представляющих собой биомолекулярные липидные слои и мозаично вкрапленные в них липопротеидопигментные комплексы, образующие фотохимически активные центры, и содержат также специальные компоненты, участвующие в транспорте электронов и образовании аденозинтрифосфата (АТФ). Часть хлоропласта, находящаяся между тилакоидами строма, содержит ферменты, катализирующие темновые реакции Ф. (например, превращение углерода, азота, серы, биосинтез углеводов и белков). В строме откладывается образуемый при Ф. крахмал. Хлоропласты имеют собственные ДНК, РНК, рибосомы, синтезирующие белки, и обладают некоторой генетической автономностью, но находятся под общим контролем ядра. фотосинтезирующие бактерии и большинство водорослей не имеют хлоропластов. Фотосинтетический аппарат большинства водорослей представлен специализированными внутриклеточными органеллами – хроматофорами, а фотосинтезирующих бактерий и сине-зелёных водорослей – тилакоидами (мембраны их содержат пигмент бактерио-хлорофилл или бактериовиридин, а также др. компоненты реакций Ф.), погруженными в периферические слои цитоплазмы.
Фаза первичных превращений и запасания энергии в процессе Ф. Воснове Ф. растений лежит окислительно-восстановительный процесс, в котором 4 электрона (и протона) поднимаются от уровня окислительно-восстановительного потенциала, соответствующего окислению воды (+ 0,8 в ) до уровня, соответствующего восстановлению CO 2с образованием углеводов (– 0,4 в ). При этом увеличение свободной энергии реакции восстановления CO 2до уровня углеводов составляет 120 ккал/моль, а суммарное уравнение Ф. выражается как:
CO 2+ H 2O 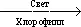 C (H 2O) + O 2+ 120 ккал/моль.
C (H 2O) + O 2+ 120 ккал/моль.
Энергия моля квантов (Эйнштейна) красной части спектра составляет около 40 ккал/моль. Т. о., для Ф., идущего в соответствии с приведённым уравнением, было бы достаточно поглощения энергии 3 квантов на молекулу CO 2(или на выделение молекулы O 2). Однако в окислительно-восстановительной реакции от воды к CO 2должны быть перенесены 4 электрона, причём перенос каждого из них осуществляется в ходе двух последовательных фотохимических реакций. Поэтому квантовый расход при оптимальных условиях составляет 8–12 квантов на молекулу O 2, а максимальная эффективность преобразования энергии красного света – около 30° %. В полевых условиях вследствие неполного поглощения света, энергетических затрат на дыхание и др. потерь, а также ограниченности вегетационного периода эффективность усвоения солнечной энергии с.-х. растениями в умеренных широтах обычно не выше 0,5–1,3%. Сопоставление этих цифр c теоретическим максимальным значением указывает на существование значительных резервов, которые могут быть использованы в будущем. Для некоторых культур с.-х. растений удаётся в специальных условиях повысить энергетическую эффективность до 5–6% и даже выше (при культивировании водорослей до 7–10%).
Ни CO 2, ни вода непосредственно не поглощают свет, посредником во взаимодействии этих соединений с квантами служит хлорофилл а, включенный в структуру хлоропласта или хроматофора и образующий функциональные фотосинтетические единицы, состоящие из нескольких сотен молекул пигмента и реакционных центров. Основная часть сопровождающих пигментов (хлорофилл b, каротиноиды, фикобилины и др. и коротковолновые формы хлорофилла а ) выполняет функцию светособирающей антенны. При поглощении квантов их молекулы переходят в возбуждённое состояние, которое путём миграции энергии передаётся на молекулу хлорофилла а, находящуюся в реакционном центре. Эффективность передачи энергии обусловлена близким расположением молекул, а также наличием нескольких агрегированных форм хлорофилла а, участвующих в формировании реакционных центров и образующих нисходящую лестницу энергетических уровней. Возможен полупроводниковый перенос электрона по агрегированному пигменту. В реакционном центре происходит основной акт Ф. – разделение зарядов с последующим образованием первичного окислителя и первичного восстановителя. Существуют два типа центров ( рис. 1 ), один из которых включен в пигментную фотосистему I (ФС I), а др. – в фотосистему II (ФС II). В фотореакции, связанной с разложением воды, участвует ФС II: пигментом её центра служит хлорофилл а с максимумом поглощения 680 нм, гипотетическим первичным восстановителем – Q (вероятно, цитохром ) , а первичным окислителем – сложный комплекс Z. Возбуждение пигментной молекулы центра P 680сопровождается разделением зарядов и образованием окисленного Z +, который участвует в окислении воды и выделении O 2. Полагают, что в систему разложения воды, пока мало изученную, входят неизвестные ферменты, ионы марганца и бикарбонат. Первичный восстановитель Q (проявляется по индукции флуоресценции) ФС II передаёт электрон переносчикам (цитохромы b, f, пластохинон, пластоцианин) фотосинтетической электронной транспортной цепи к реакционному центру ФС I. Пигментом этого центра служит хлорофилл а с максимумом поглощения 700 нм, первичным восстановителем – неидентифицированное вещество Х. Восстановленный Х передаёт электрон ферредоксину – железосодержащему белку, который восстанавливает никотинамидадениндинуклеотидфосфат (НАДФ). Его восстановленная форма – НАДФ-Н запасает основную часть энергии света. Др. часть энергии электронного потока запасается в виде АТФ (фотосинтетическое фосфорилирование), который образуется на нисходящем участке переноса электронов между ФС II и ФС I (нециклическое фотофосфорилирование) или при круговом замыкании потока в ФС I (циклическое фотофосфорилирование). Фосфорилирование, возможно, происходит по хемиосмотическому механизму за счет электрического потенциала и градиента концентрации Н +, возникающих при индуцировании светом электронного потока в мембранных структурах тилакоидов. Экспериментально обнаружено, что освещение индуцирует электрический потенциал на мембране хлоропласта. Описанное последовательное соединение двух фотореакций I и II наиболее вероятно, хотя обсуждается возможность параллельного соединения реакций. Предполагают, что фотосинтезирующие бактерии осуществляют Ф. с участием лишь одной пигментной фотосистемы, однако этот вопрос нельзя считать решенным. Фотофизические и фотохимические стадии заканчиваются за 10 -12–10 -8 сек разделением зарядов и последующим образованием первичного окислителя и восстановителя. Границей первичных биофизических и биохимических процессов обычно считают появление первых химически стабильных продуктов – НАДФ-Н и АТФ. Эти вещества («восстановительная сила») используются затем в темповых процессах восстановления CO 2.
Читать дальшеИнтервал:
Закладка: