Дмитрий Сахаров - Генеалогия нейронов

- Название:Генеалогия нейронов
- Автор:
- Жанр:
- Издательство:Издательство «Наука»
- Год:1974
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Дмитрий Сахаров - Генеалогия нейронов краткое содержание
Д. А. САХАРОВ
ГЕНЕАЛОГИЯ НЕЙРОНОВ
ИЗДАТЕЛЬСТВО «НАУКА» МОСКВА 1974
АКАДЕМИЯ НАУК СССР ИНСТИТУТ БИОЛОГИИ РАЗВИТИЯ
© Издательство «Наука», 1974 г.
Генеалогия нейронов - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
На рис. 6 суммированы результаты, касающиеся знака ответа на ацетилхолин нейронов, находящихся в разных ганглиях подглоточного комплекса. Данные, относящиеся к картированным индивидуальным нейронам, на этих схемах не отражены, они изложены в разделе 4.2.5. Рисунки дают представление о региональных особенностях ответов нейронов на ацетилхолин. Так, на вентральной поверхности педальных ганглиев и в той области правого париетального ганглия, где расположены крупные нейросекреторные клетки (группа D), ацетилхолин почти без исключений вызывает гиперполяризацию. Противоположные, деполяризующие эффекты наблюдаются, как правило, на дорзальной поверхности висцерального ганглия в зоне, занятой преимущественно нейронами группы F. Несколько более пёстрые результаты, полученные в аналогичных условиях голландскими авторами, которые изучали ответы нейронов, расположенных на дорзальной поверхности комплекса [346], объясняются, по всей вероятности, тем, что этими авторами при оценке знака ответа не выдерживалось отмеченное выше стандартизирующее условие.
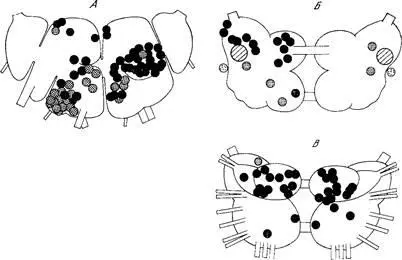
Рис. 6. Знак ответа на ацетилхолин со стороны нейронов, расположенных в разных участках ЦНС виноградной улитки.
А — ганглии висцеральной дуги; Б, В — педальные ганглии с дорзальной и вентральной стороны. На схемах суммированы результаты разных опытов, но данные, полученные на индивидуально идентифицируемых клетках, в рисунок не включены и сообщаются в тексте (то же относится к рис. 7). Ионофоретическая аппликация. Заштрихованный кружок указывает позицию клетки, деполяризуемой медиатором, чёрный кружок — гиперполяризуемой клетки.
Об особом, «двойном» действии ацетилхолина на некоторые нейроны будет сказано ниже, при описании группы G ( 4.2.5.).
Ионные механизмы эффектов ацетилхолина были нами исследованы в нескольких случаях. Деполяризующие эффекты оказались натрийзависимыми и гиперполяризующие — зависимыми от ионов хлора [24]. Пока не обнаруживались другие типы эффектов, найденные на нейронах других моллюсков.
В литературе имеются данные о том, что холинорецепторы, ответственные за разные ионные эффекты ацетилхолина на нейроны гастропод, имеют разное строение, что выражается их отношением к холинолитикам и холиномиметикам [208, 232].
Первичные катехоламины.Признано, что дофамин, подобно ацетилхолину, выполняет медиаторные функции в нервной системе гастропод. Как правило, чувствительные к дофамину нейроны моллюсков, в частности аплизии, отвечают на него гиперполяризацией, но бывают и противоположные эффекты [83]. Глайзнер, по-видимому, впервые нашел у садовой улитки клетку, реагирующую на дофамин деполяризацией [170]. У виноградной улитки нам удалось исследовать в этом отношении только поверхностные нейроны задне-медиальной области правого париетального ганглия (дорзальная клеточная кора). В этой области большинство клеток, в частности нейроны ППа1, ППа2 и клетки группы D, гиперполяризуются и тормозятся дофамином, но имеется также несколько небольших клеток, отвечающих на дофамин деполяризацией.
Чувствительностью к норадреналину обладают, как правило, те же нейроны садовой улитки, которые реагируют на дофамин [170]; для виноградной улитки собственными данными по действию норадреналина мы не располагаем.
Серотонин.В литературе детальные данные о клеточных эффектах серотонина имеются только для садовой улитки Helix (Cryptomphallus) aspersa. Сначала Гершенфельду и Стефани удалось заметить только возбуждающие эффекты, которые развивались с большой латентностью, из чего авторы сделали вывод, что серотонин действует на расстоянии от тела нейрона [168]. Позже Глайзнер сообщил, что у этого вида имеются также нейроны, отвечающие на серотонин торможением [170], а Гершенфельд нашел, что тормозные эффекты имеют разную ионную и рецепторную природу в правом и левом париетальном ганглиях [165]. Все эти исследования проводились на дорзальной поверхности ганглиев подглоточного комплекса.
У виноградной улитки мы с Я. Шаланки также описали сначала только возбуждающие эффекты серотонина [283]. В ходе дальнейшей работы мы с Г. Н. Коробцовым нашли в этой и других областях ЦИС виноградной улитки разные типы ответов на ионофоретически апплицируемый серотонин [25]. Отдельные клетки, отвечающие на серотонин торможением, были обнаружены в левом и правом париетальных ганглиях, т. е. там, где у садовой улитки их нашел Гершенфельд.
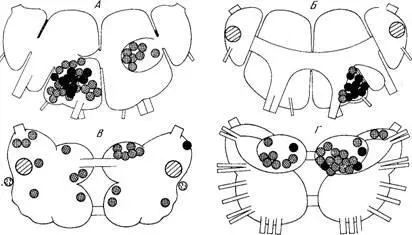
Рис. 7. Знак ответа на серотонин со стороны нейронов, расположенных в разных участках ЦНС виноградной улитки.
А ,Б — ганглии висцеральной дуги; В, Г — педальные ганглии с дорзальной и вентральной стороны соответственно. Условия те же, что на рис. 6.
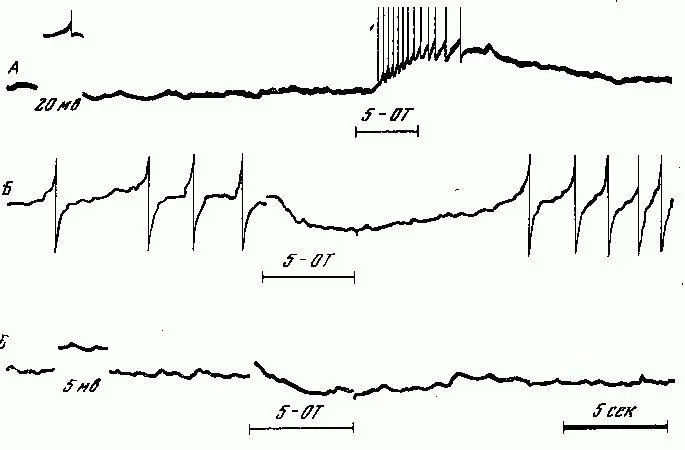
Рис. 8. Противоположные по знаку ответы клеток висцерального ганглия (дорзальная поверхность) на серотонин (ионофоретическая аппликация).
А — клетка, деполяризуемая серотонином; Б — клетка, гиперполяризуемая серотонином (остановка спонтанной активности); В — та же клетка при более негативном значении мембранного потенциала: генерация отсутствует, серотонин гиперполяризует.
Кроме того, в висцеральном ганглии найдена большая, компактно расположенная группа клеток, у которых серотонин тоже вызывает гиперполяризацию. Эта группа (группа G) хорошо представлена на вентральной поверхности ганглия, но выходит и на дорзальную поверхность. Результаты ионофоретической аппликации серотонина на нейроны этого ганглия схематически подытожены на рис. 7, а на рис. 8 представлены записи эффектов серотонина на клетки ганглия. В педальном ганглии эффекты серотонина оказались в основном деполяризующими, но и здесь были обнаружены отдельные нейроны, отвечающие гиперполяризацией (рис. 7, Г). Ионный механизм был изучен только на тех клетках, которые отвечают на серотонин деполяризацией: эффект всегда оказывался натрийзависимым. Интересно отметить, что это относится и к гигантским метацеребральным нейронам, которые хорошо отвечают на серотонин несмотря на то, что сами являются серотонинергическими (см. 4.2.5). Подробнее о действии серотонина на идентифицированные клетки см. ниже. Здесь отметим лишь, что стимулирующие эффекты серотонина, вероятно, могут иметь разные механизмы. По нашим наблюдениям, одни нейроны отвечают на серотонин быстро развивающейся деполяризацией, которая прекращается вслед за прекращением аппликации серотонина. Таковы, например, гигантские метацеребральные нейроны (клетки ПЦ1 и ЛЦ1 — см. 4.2.5.). В других случаях стимуляция характеризуется большой латентностью, длительным последействием и как бы избирательно направлена на осциллогенез; такое действие серотонина нельзя имитировать деполяризующим током. Этот тип эффекта наблюдается у нейронов пептидергического типа — группы А, В, D, крупные белые клетки ганглиев висцеральной дуги [26]; он выражен и у некоторых педальных нейронов, которые, по-видимому, не являются пептидергическими.
Читать дальшеИнтервал:
Закладка:









