Александр Зубов - Становление и первоначальное расселение рода «Homo»

- Название:Становление и первоначальное расселение рода «Homo»
- Автор:
- Жанр:
- Издательство:Алетейя
- Год:2011
- Город:Санкт-Петербург
- ISBN:978-5-91419-371-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Зубов - Становление и первоначальное расселение рода «Homo» краткое содержание
В частности, глобальный анализ локализации и датировок палеоантропологических находок и местонахождений древнего человека дает возможность с гораздо большей точностью, чем ранее, установить место и время возникновения первых человеческих популяций («колыбель» рода Homo), а также составить представление о поведении и образе жизни первых людей. Новое слово может быть сказано и о таком важнейшем феномене человеческой эволюции, как миграции и связанные с ними процессы адаптации к меняющейся среде, т. е. становится возможным лучше понять те биологические, социальные и поведенческие факторы, которые позволили роду Homo заселить и освоить практически всю территорию нашей планеты. Становление древнейшего человека и его опыт как «первооткрывателя» всех земель ойкумены — центральная тема данной книги. Монография рассчитана на широкий круг читателей.
Становление и первоначальное расселение рода «Homo» - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Этапы становления Homo erectus/ergaster хорошо прослеживаются на краниологических материалах, относящихся к периоду 2+ — 1,5 млн. лет до современности. На это время приходится целый ряд находок «промежуточного» характера, вызывающих постоянные споры по поводу их систематического положения (рис. 6 a-d). Древнейшим звеном мы в данном случае будем считать KNM-ER 1470, который сейчас относят к Homo rudolfensis , т. е. к эволюционному уровню хабилиса, хотя в его морфологических особенностях исследователи всегда отмечали присутствие черт Homo erectus (рис. 6 а). Здесь перед нами — фактически уже знакомая проблема таксономического диагноза древнейших Homo , на деле являющихся ранней формой Homo erectus, которую мы определили как Homo habilis pre-erectus. — предшественника H. erectus. Ее полный морфологический комплекс проявился в черепной серии из Дманиси. Поэтому нет необходимости возводить № 1470 в ранг отдельного вида, что было сделано в то время, когда еще не было ключевого материала для выделения раннего подвида Homo erectus. Этот экземпляр, по нашему мнению, должен быть включен вместе с находками в Дманиси и южноафриканскими черепами SK 847 и Stw 53 в подвид Homo habilis pre-erectus, «типовым» экземпляром которого является череп D2700 из Дманиси (рис. 6 b). Часть древней популяции ранних Homo продолжала эволюционировать на территории Кении и Эфиопии, где примерно 1,6 млн. лет назад выделился вариант KNM-WT 15000 — Нариокотоме III (рис. 6 с), отличающийся определенным своеобразием, особенно в том, что касается посткраниального скелета (большая длина тела). Иногда эту находку прямо относят к Homo ergaster, но достаточно посмотреть на череп KNM-WT 15000 в латеральной норме, чтобы убедиться в его очевидном сходстве с Homo hablis pre-erectus. Это сходство проявляется также в морфологии зубов: уникальная форма верхних резцов, коронка которых сильно расширяется в сторону режущего края отмечается на черепах Нариокотоме и Дманиси D2700; очень большая ширина тригонида по отношению к талониду характерна для первых нижних моляров Нариокотоме и D2735. Речь здесь идет о редких признаках, что наводит на мысль об общих генетических корнях палеодема Дманиси и ранних африканских Homo. Эволюционный ряд краниологических типов от истоков рода Homo до дефинитивных форм Homo erectus/ drgaster завершает «типовой» для Homo ergaster череп KNM-ER 3733 из Кении (рис. 6 d). По объему мозговой коробки (848 см 3) этот экземпляр мало отличается от Нариокотоме (880 см 3), но свод черепа у него выше, а чешуя лобной кости, отделенная от надглазничного валика глубокой канавкой, гораздо менее наклонна, чем на черепах Homo habilis pre-erectus. Граница между этими таксонами определяется не только по данному признаку: в Рассмотренной работе о черепах из Дманиси ( Rightmi-er et al., 2006 ) авторы провели сравнительное исследование черепов ранних гоминид по параметрам строения нёба, причем был построен бивариантный график, на координатах которого отложены глубина нёба и нёбный индекс ( р. 132 ). Точки, обозначающие черепа KNM-ER 3733 и D2700 расположились на поле графика на очень значительном расстоянии друг от друга. Перед нами наглядная картина становления нового таксона африканских Homo, который многие исследователи сейчас определяют как вид Homo ergaster, но автор данной статьи в интересах последовательности своей позиции в систематике гоминин предпочитает рассматривать термин «ergaster» скорее как подвидовой , входящий в триномен Homo erectus ergaster. Это наименование, кстати, совместимо с позицией тех антропологов, которые предпочитают не противопоставлять эволюционный уровень форм ergaster и erectus, считая биномен Homo erectus, универсальным для африканских и азиатских гоминин, относящихся к стадии «первых настоящих людей». В цитируемой статье, посвященной человеку из Дманиси, это проявляется
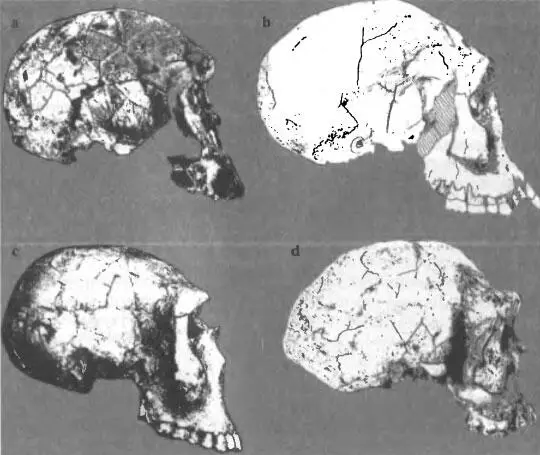
Эволюция черепа Homo erectus / ergaster от древнейших форм (KNM-ER 1470) (а) через «типовой» вариант» Н. habilis pre-erectus (b) (Дманиси D2700), промежуточный тип Нариокотоме III (с) — до «классического» Homo erectus ergaster KNM-ER 3733 (d)
вполне отчетливо: на стр. 133 читаем «…African erectus (= ergaster)». Подвидовой уровень различий между африканскими и азиатскими Н. erectus авторы связывают с большой, по их мнению, скоростью первоначального расселения ранних Homo после «исхода» из Африки и недостатком времени для возникновения существенных таксономических различий (« …there would have been little time for the diverging groups to develop distinctive characters» ( « …дивергирующие группы имели, вероятно, мало времени чтобы выработать отличительные признаки». (op. cit. р. 139 ). Выше приводились рассуждения, согласно которым незначительная интенсивность процессов дивергенции и видообразования у человека может быть объяснена на основании специфических особенностей рода Homo. Все же пока сходство африканских и азиатских эректусов на подвидовом уровне не будет твердо доказано (а у меня остаются некоторые сомнения на этот счет), «не следует недооценивать определенных различий между сравниваемыми формами» ( Васильев, 1999 ).
В итоге, генеалогическое древо рода Homo может выглядеть, по нашим представлениям, следующим образом (рис. 7). Род Homo выделяется из среды австралопитековых как Homo habilis sensu lato около 2,5 млн. лет назад и через 200–300 тыс. лет происходит разделение этого таксона на две ветви: Homo habilis habilis et Homo habilis pre-erectus. Последний широко расселяется го территории Африки и осуществляет «исход» — на земли Евразии, сравнительно быстро (?) достигая Юго-Восточной и Восточной Азии, сохранив при этом видовую ( erectus ) и, отчасти, подвидовую модель ( mojokertensis ~ pre-erectus ). Далее происходит дивергенция подвида Н. erectus erectus и его восточноазиатского ответвления — Н. erectus pekinensis. Лантьянслого эректуса, очевидно, можно отнести к Н. habilis pre-erectus ( mojokerlensis ),
В Африке анагенегическая эволюция приводит к становлению Н. erectus ergaster, который может рассматриваться как «opposite number» азиатского Homo erectus erectus. От него происходят Н. erectus mauretanicus и Н. antecessor (=ранний H. heidelbergensis ).
Возникновение нового таксона, достигшего максимально совершенного уровня адаптации в данных условиях, часто связано со «взрывами» миграционной активности. Во всяком случае, на протяжении всей линии эволюции рода Homo эта закономерность проявляется на самых различных уровнях.
Существует мнение, согласно которому граница Африканского континента в палеолите пересекалась не только несколько раз в направлении «out-of Africa», но также были возможны миграции противоположного направления — «back-to-Africa» (Wood & Turner, 1995 ). Аргументация этой точки зрения строится на фактах наличия, по мнению некоторых антропологов, морфологических черт Н. erectus sensu stricto на черепах, по времени относящихся к периоду 0,9–1,2 млн. лет до современности, найденных на территориях Европы (Чепрано) и восточной Африки (ОН 9). Б. Вуд и А. Тернер предположили, что древнейшие Homo вышли из Африки в позднем плиоцене, а Н. erectus эволюционировал в Азии, но потом вернулся в Африку (Wood & Turner, 1995 ). Антропологи и археологи последующих лет в основном положительно восприняли идею возможности возникновения Н. erectus в Азии ( Hyodo et al., 2002, Деревянко, 2006 ). Исследователи справедливо замечают при этом, что признание этой гипотезы в сильной степени зависит от даты первоначального появления Homo в Азии, а, надо сказать, что в этом вопросе существует довольно широкий спектр мнений. Если исходить из наиболее распространенной сейчас датировки ( Swisher et al., 1994 ), то «сценарий» Б. Вуда и А. Тернера представляется вполне правдоподобным: человек пришел в Индонезию на ранней стадии еще не сформировавшегося вида Н. erectus и эволюционировал как Н. erectus erectus, в то время как в Африке развился вариант Н. erectus ergaster. Однако, гипотеза «back-to-Africa » пока не получила широкого распространения среди специалистов. Представляется мало понятным «поворот на 180°» самой дальней
Читать дальшеИнтервал:
Закладка:









