Максим Франк-Каменецкий - Самая главная молекула. От структуры ДНК к биомедицине XXI века

- Название:Самая главная молекула. От структуры ДНК к биомедицине XXI века
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2017
- Город:Москва
- ISBN:978-5-9614-4522-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Максим Франк-Каменецкий - Самая главная молекула. От структуры ДНК к биомедицине XXI века краткое содержание
Самая главная молекула. От структуры ДНК к биомедицине XXI века - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Однако особого интереса открытие не вызвало. Мало ли какой бывает ДНК в вирусах! Иногда она находится там в виде одной из двух комплементарных цепей. Порой эта цепь замкнута в кольцо. Но заведомо известно и много случаев, когда внутри вирусной частицы ДНК линейна.
Все же поиск кольцевых ДНК продолжался. И постепенно выяснилось, что даже в тех случаях, когда ДНК в вирусной частице линейна, она, как правило, замыкается в кольцо после проникновения вируса в клетку. Оказалось, что перед началом репликации такая линейная молекула переходит в форму (ее называют репликативной), в которой обе комплементарные цепи ДНК образуют замкнутые кольца (рис. 26). Кольцевыми оказались ДНК бактерий, в частности, кишечной палочки. Плазмиды, эти незаменимые в генной инженерии переносчики генов, всегда кольцевые. Короче, трудно назвать случаи, когда ДНК работает в прокариотической клетке, не находясь в кольцевом состоянии. Отметим сразу, что у эукариот хромосомная ДНК всегда линейна. Мы еще вернемся к этому фундаментальному различию между двумя основными царствами живых существ. А пока сосредоточим свое внимание на прокариотах. Зачем прокариотической клетке замыкать молекулы ДНК в кольца? Что это дает? К каким изменениям свойств молекул приводит? Чтобы ответить на эти вопросы, надо было подробно изучить эту новую форму ДНК.

Рис. 26.В замкнутой кольцевой ДНК две комплементарные цепочки образуют зацепление высокого порядка
Сверхспирализация и топоизомеразы
Для нас сейчас важнее всего то, что в молекуле ДНК комплементарные цепи обвивают друг друга подобно двум лианам, и когда каждую из цепей замыкают, то два кольца оказываются зацепленными так, что их невозможно развести. Простейшее зацепление двух колец известно всем – это символ бракосочетания (рис. 27). Только две комплементарные цепи в ДНК сцеплены друг с другом гораздо сильнее.
Количественно степень зацепленности двух колец характеризуется величиной, называемой порядком зацепления и обозначаемой Lk (от английского слова linking). Определить эту величину для любого зацепления очень легко. Нужно представить себе, что на одно кольцо натянута мыльная пленка, и подсчитать, сколько раз второе кольцо протыкает эту пленку. Тогда легко убедиться, что для символа бракосочетания Lk = 1, а для зацепления, изображенного на рис. 26, Lk = 9.
Величина Lk замечательна тем, что ее значение для заданной пары колец не может измениться, как бы мы ни гнули эти кольца, лишь бы не рвали их. Поэтому математики говорят, что Lk есть топологический инвариант системы, состоящей из пары колец. А без помощи математиков молекулярным биологам никогда не удалось бы разобраться в свойствах кольцевых ДНК.
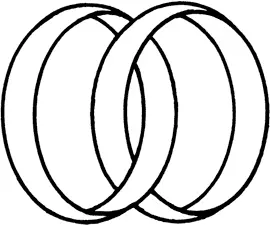
Рис. 27.Простейшее зацепление – символ бракосочетания
Итак, если мы превратили ДНК в кольцевую замкнутую молекулу, то созданный в ней порядок зацепления двух цепей не может измениться, что бы мы ни делали с молекулой, пока сахарофосфатные цепи, образующие «хребет» каждой из комплементарных цепочек, остаются целыми и невредимыми. Благодаря этому обстоятельству замкнутые кольцевые (зк) ДНК обладают совершенно особыми свойствами, резко отличающими их от линейных молекул. Самое главное заключается в том, что в зкДНК может быть запасена впрок энергия в виде так называемых сверхвитков.
Чтобы пояснить только что сказанное, представим линейную ДНК в каких-то определенных внешних условиях. В такой ДНК на один виток двойной спирали приходится вполне определенное число пар оснований. Это величина γ 0. В двойной спирали Уотсона—Крика γ 0= 10, но она может немного меняться (всего лишь на десятые доли, но сейчас для нас это важно) при изменении внешних условий. Допустим теперь, что из линейной молекулы сделали кольцевую, прибегнув к минимальному насилию. Проще всего представить себе, что мы превратили молекулу в окружность и «заклеили» концы каждой из цепей. Чему будет равно Lk ? Ясно, что Lk = N / γ 0, где N – число пар оснований в молекуле.
Теперь изменим внешние условия. Молекула ДНК приобретает другое равновесное значение числа пар оснований на виток – γ 0, хотя величина Lk измениться не может. Что же происходит? Молекула стремится обрести положенный порядок зацепления: Lk' = N / γ 0, но не в состоянии себе этого позволить, ей уже навязано иное значение Lk. Подобное случается и с брачными узами. Когда они заключались, то Lk = 1, но вот условия изменились, той или другой стороне хочется расторгнуть брак, т. е. сделать Lk равным нулю. Возникает очень напряженная обстановка. Нечто похожее происходит и с ДНК. Молекула оказывается в напряженном, энергетически невыгодном состоянии сверхспирализации.
Обычно сверхспирализованные молекулы принимают форму, показанную на рис. 28. Количественно сверхспирализация характеризуется величиной τ = Lk-N / γ 0. Подобно тому, как самой двойной спирали приписывается определенный знак (положительный для правой спирали и отрицательный для левой), так и сверхспирализация может в принципе быть положительной или отрицательной. На рис. 28 двойная спираль правая, как и положено для ДНК, а сверхспирализация отрицательна.
Последнее утверждение может вызвать недоумение. Ведь кажется, что сверхспираль на рис. 28 правая, а не левая. Это один из парадоксов, с которыми приходится сталкиваться при изучении сверхспирализации. Чтобы проще было во всем этом разобраться, возьмите кусок резинового шланга длиной чуть меньше метра, по возможности жесткого. Вставьте в один конец шланга какой-нибудь штырь так, чтобы он немного торчал наружу и на него можно было надеть другой конец, замкнув шланг в кольцо. Важно, что концы шланга после замыкания не должны свободно прокручиваться относительно друг друга.
Теперь можно моделировать сверхспирализацию. Для этого, держа один конец неподвижным, вращайте другой конец шланга вокруг оси штыря так, чтобы ось шланга образовала левую винтовую линию. Затем дайте замкнутому в кольцо шлангу принять наиболее выгодное для него положение, придерживая его двумя пальцами одной руки. Вы убедитесь, что он примет форму, аналогичную изображенной на рис. 28.
Рис. 28.Такой вид принимает сверхспиральная ДНК. Сверхспирализация отрицательная
По мере того как из клеток аккуратно выделяли все новые ДНК и определяли их состояние, вновь и вновь убеждались в том, что эти ДНК не только замкнуты в кольцо, но и завиты в сверхвитки; при этом сверхспирализация абсолютно во всех случаях оказывалась отрицательной. Стало ясно, что сверхспирализованное состояние ДНК не исключение, как думали вначале, а правило. Но тут возникло сомнение – а такова ли ДНК там, внутри клетки? Пришлось признать, что, скорее всего, нет, не такова. По-видимому, сверхспирализация – это реакция на насильственное извлечение ДНК из родной стихии, ведь условия, в которых пребывает ДНК внутри клетки, конечно же, отличаются от условий после ее извлечения.
Читать дальшеИнтервал:
Закладка:









