Сергей Савельев - Происхождение мозга

- Название:Происхождение мозга
- Автор:
- Жанр:
- Издательство:Веди
- Год:2005
- ISBN:5-94624-025-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Сергей Савельев - Происхождение мозга краткое содержание
Происхождение мозга - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Ещё более парадоксальная ситуация возникает при анализе слуховой системы рипидистий. По общему мнению, лабиринт у этих животных развит намного лучше, чем у их потомков. Полукружных каналов три и они очень крупные. У примитивных амфибий это также хорошо развитый орган. Он включает в себя три органа чувств: слуховой аппарат (или его кожный аналог — боковая линия), рецептор линейного ускорения (гравитационный рецептор) и рецептор углового ускорения (и торможения). Вполне справедлив вопрос о том, зачем такая совершенная система управления движением, если животное ползает в мелком водоёме или грязи миллионы лет. Если бы это было действительно так, то один из полукружных каналов редуцировался, как и у круглоротых. Поскольку этого не произошло, можно с уверенностью сказать, что обладатели такого развитого рецептора ускорения существовали в трёхмерной среде, а не барахтались в пересыхающих лужах. С этим выводом согласуются особенности строения специфической сенсорной системы, характерной для всех саркоптеригий. Рецепторы этой системы были расположены в поверхностных слоях покровного скелета. Под эмалеподобной поверхностью у древних рипидистий и двояко-дышащих располагается своеобразный дентин с системой поровых каналов и тонкими сенсорными порами. Найденные у рипидистий образования сходны по строению с ампулами Лоренцини и электрорецепторами современных хрящевых и костистых рыб (Thomson, 1975; Vorobyeva, 1980). Эти органы необходимы для ориентации в темноте, где другие дистантные рецепторы малоэффективны. Акулы и костистые рыбы используют этот рецепторный орган на больших глубинах или в толще мутной воды. Учитывая совершенство морфологического строения ампулярного аппарата рипидистий, кажется вполне логичным, что порог их чувствительности был близок к порогу чувствительности отдельного рецептора в ампуле Лоренцини современных акул. Если даже его чувствительность была в несколько десятков раз ниже, это всё равно был очень совершенный орган, который позволял ориентироваться и искать добычу по искажениям электрического поля. Следовательно, саркоптеригий сформировали специализированную систему рецепции, для которой нужен довольно большой объём воды.
В неглубоких и пересыхающих водоёмах такой дистантный анализатор практически бесполезен. По сути дела создаётся взаимоисключающая ситуация: наиболее совершенный рецептор возник и усовершенствовался в условиях, где его невозможно использовать. По-видимому, этот орган сформировался и использовался Crossopterygii в более подходящих условиях, чем пересыхающие водоёмы.
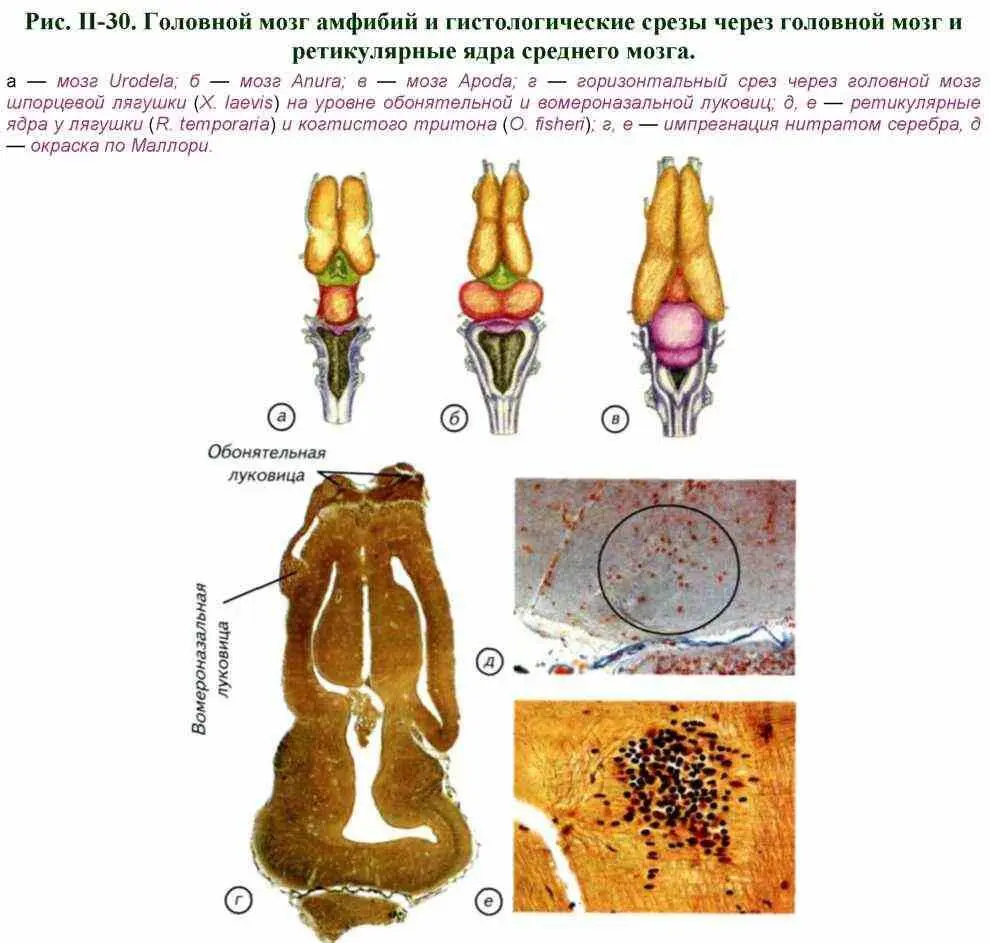
Довольно своеобразно выглядит организация зрительной системы саркоптеригий. У них обнаружены небольшие глазницы, которые свидетельствуют о незначительном развитии зрительной системы (рис II-31). Это странная ситуация, поскольку в «мелководных и пересыхающих» водоёмах зрение должно играть доминирующую роль. Однако вопреки всем ожиданиям развитая зрительная система у рипидистий не встречается. Среди разлагающихся органических остатков абсолютно «ненужная» обонятельная система быстро эволюционирует и дифференцируется в две системы с самостоятельной иннервацией и соответствующими центрами в головном мозге. Возникли две системы водно-воздушного обоняния. У амфибий наряду с основным органом обоняния появилась добавочная — вомероназальная (якобсонова) система дистантной хеморецепции. Таким образом, обоняние интенсивно эволюционировало, приспосабливалось к воздушной среде, а зрительная система изменялась незначительно.
Попробуем представить себе загадочного предка наземных позвоночных в широко признанной среде
обитания. Неглубокий, прогретый и периодически пересыхающий водоём так заполнен разлагающимися органическими остатками, что у рипидистий возникли лёгкие для воздушного дыхания. В этом водоёме при недостатке пищи находится слабовидящее животное, лишённое возможности использовать развитое обоняние, и практически глухое. Эту безрадостную картину дополняет ненужный в мелких водоёмах, но хорошо развитый рецептор ускорения и такой же никчёмный, но совершенный дистантный электрорецептор. Животное обладает всем, что ему совершено не нужно, и не имеет того, что ему просто необходимо. Все перечисленные противоречия приносятся в жертву одной идее — мелководной промежуточной среде обитания, которая считается «естественным» этапом между водной средой и сушей.
Таким образом, перечисленные особенности строения Crossopterygii ставят под сомнение гипотезу «прибрежно-мелководного» происхождения наземных позвоночных. Вероятность «счастливой случайности», позволившей преодолеть эти проблемы, крайне мала.

Следовательно, если мы хотим ясно представить события, приведшие к появлению тетрапод, необходимо ответить на ряд простых вопросов. Почему предки позвоночных стали жить у побережья? Отчего возникла необходимость в воздушном дыхании? Почему слаборазвитые конечности оказались более эффективными, чем работа совершенной туловищной мускулатуры? Почему у примитивных четвероногих хорошо развиты обоняние, системы контроля движения и плохо развито зрение?
Ответы на эти вопросы легко получить, если перечислить биологические условия, для которых известная организация органов чувств и мускулатуры Crossopterygii и дошедших до нас останков древних амфибий (см. рис. II-30; II-31) была бы оптимальной. Водное, как и воздушное, обоняние имеет биологический смысл, если оно пригодно для поиска пищи, ориентации и размножения. Нам известно, что у амфибий впервые сформировалась добавочная обонятельная система — якобсонов орган. В результате амфибиям стала доступна для химического анализа как воздушная, так и водная среда. Столь глубокие изменения органов обоняния, а не зрения, говорят о том, что зрительный анализатор был явно менее эффективным в тот период, хотя, казалось бы, в пересыхающих прибрежных водоёмах должно было бы преобладать зрение. Следовательно, выход на сушу осуществлялся не через мелководные водоёмы, а в какой-то другой переходной среде.
§ 33. Выход амфибий на сушу
Наиболее вероятным биотопом перехода из воды на сушу для кистепёрых были прибрежные водновоздушные лабиринты (рис. II-32; II-33). В них находились как морская, так и стекающая с берега пресная вода, полузаполненные воздухом и водой многочисленные камеры и связанные между собой полости. Это могло быть размытое устье реки или набор каверн, возникших по геологическим причинам. Подобные каверны широко распространены на Тихоокеанском побережье Южной Америки, на восточном побережье Африки и в Индокитае. Подходящей переходной средой для выхода на сушу являются почвенные каверны на границе мангровых зарослей или пещеристые берега заболоченных районов современного Цейлона. Ещё одной причиной появления многочисленных районов с полуводными лабиринтами могли стать растения. В период появления наземных позвоночных у большей части растений были полые стволы. Собственно говоря, почти во всех растениях этого позднего периода были полости различных размеров. Растения с полостями внутри погибали, но не разрушались, что позднее создало каменноугольные залежи и даёт представление о масштабах процесса. Толщина каменноугольных пластов в настоящее время достигает десятков метров, они распространены на огромных территориях.
Читать дальшеИнтервал:
Закладка:



