Сергей Савельев - Происхождение мозга

- Название:Происхождение мозга
- Автор:
- Жанр:
- Издательство:Веди
- Год:2005
- ISBN:5-94624-025-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Сергей Савельев - Происхождение мозга краткое содержание
Происхождение мозга - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
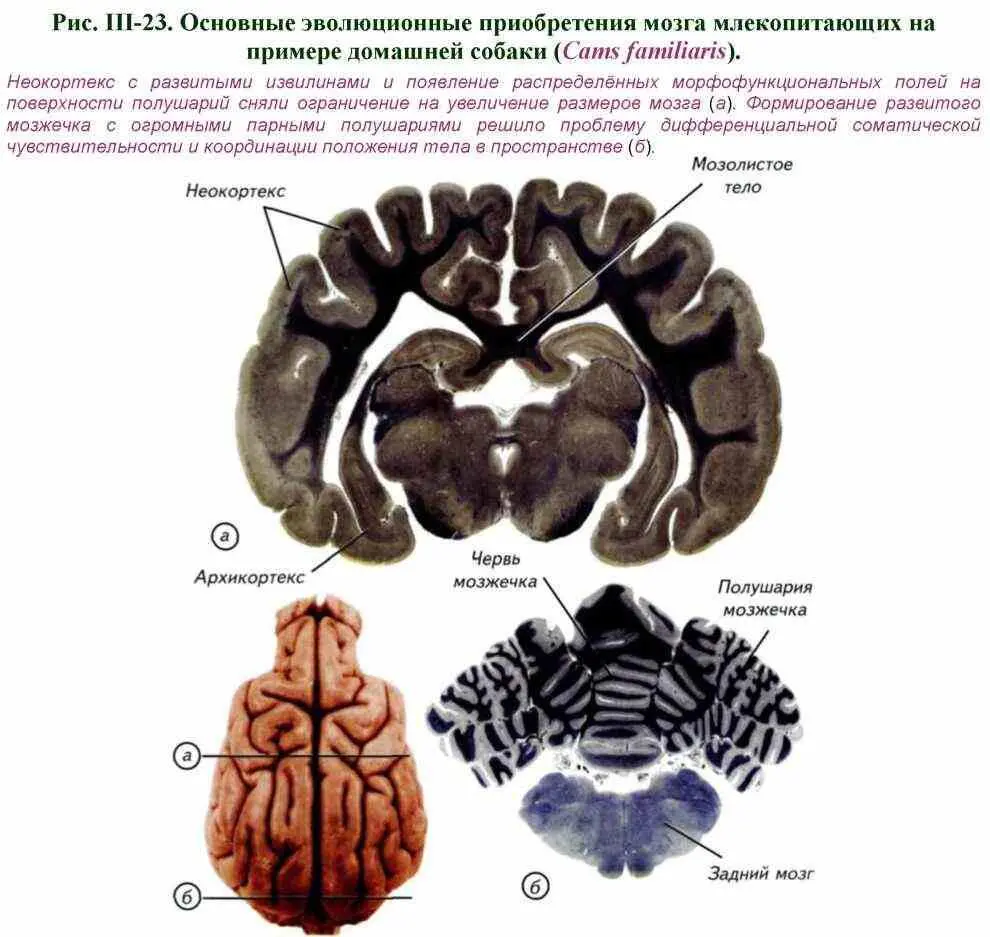
Основным отличием строения мозга млекопитающих от мозга рептилий и птиц являются крупные парные полушария переднего мозга и мозжечка (см. рис. III-18; III-19). Поверхность полушарий переднего мозга млекопитающих может быть гладкой — лиссэнцефальной или складчатой — с бороздами и извилинами. Полушария переднего мозга млекопитающих разрастаются вбок и кзади, покрывая собой промежуточный мозг, часто также средний мозг, а у приматов и китообразных — продолговатый мозг (см. рис. III-19; III-22).
Полушария переднего мозга имеют неправильную овальную форму и более или менее расширены сзади. У многих млекопитающих самых разнообразных групп (насекомоядные, грызуны, зайцеобразные) полушария гладкие, без борозд и извилин. У большинства высших млекопитающих в коре формируются борозды и извилины (см. рис. III-19-III-21). Однако извилины бывают развиты в различной степени даже в пределах одного отряда. Мозг низших приматов практически лишён борозд и извилин, тогда как у высших приматов они сходны с извилинами мозга человека (см. рис. III-22). С увеличением размеров полушарий непосредственно связано появление мозолистого тела — обширной зоны волокон, соединяющих левое и правое полушария. Это образование характерно для мозга высших млекопитающих, но отсутствует в мозге однопроходных и сумчатых (см. рис. III-18-III-21).
Полушария переднего мозга млекопитающих в процессе эволюции обычно значительно увеличивались в рострокаудальном направлении и немного — в латеральном, поэтому головной мозг большинства млекопитающих вытянут вдоль тела. Единственным исключением является организация мозга у китообразных. Это связано с тем, что у большинства китообразных передний мозг увеличивался в латеральном направлении. В результате этих особенностей эволюции у китообразных оказались особенно развитыми височные доли переднего мозга, что привело к нарушению консервативной схемы расположения полей. Зрительная область, у млекопитающих всегда расположенная на затылочном полюсе мозга, оказалась в положении теменной доли, а соматосенсорные поля сместились в лобную долю (Савельев, 1996, 2001).
У китообразных борозды и извилины достигли максимального развития (см. рис. III-19, г; III-22, б). При равной массе тела человека и дельфина мозговых борозд у последнего в 2 раза больше, чем у человека. Большое количество борозд и извилин является следствием увеличения площади поверхности коры. Причиной таких изменений может быть простое увеличение мышечной массы животного. У крупных животных значительная часть коры переднего мозга занята сенсомоторными и моторными полями, тогда как ассоциативные и сенсорные поля увеличиваются почти пропорционально размеру мозга, поэтому крупный мозг китообразных и хоботных отнюдь не свидетельствует о высоком интеллекте его обладателей.
Крупные полушария млекопитающих исторически возникли из боковой части стенки переднего мозга рептилий в результате разрастания поверхностного скопления клеток, которое носит название плаща, или коры. Небольшое скопление клеток на латеральной стенке мозга рептилий (см. рис. III-7, а), стало источником невероятного неврологического и интеллектуального прогресса млекопитающих (Филимонов, 1963, 1974). Эта однородная полоска нейронов примордиального неокортекса рептилий трансформировалась в новую кору млекопитающих. Она состоит из 6 чётко выделяемых слоёв, толщина которых у высших приматов доходит до 100 клеток (см. рис. III-20-III-22). В коре млекопитающих продублированы почти все основные функции нервной системы, которые присутствуют на уровне заднего, среднего, промежуточного и переднего мозга.
При анализе развития переднего мозга млекопитающих необходимо учитывать не только его внешнее строение, но и пространственное распределение нейронов в коре — цитоархитектоническую организацию. Кора переднего мозга млекопитающих имеет сходное строение со слоистой крышей среднего мозга низших позвоночных. Неокортекс млекопитающих имеет шестислойное строение. У взрослых млекопитающих различных видов кора может значительно различаться как по толщине, так и по выраженности отдельных слоёв (см. рис. III-22). Известно, что у животных с лиссэнцефальным мозгом поперечник коры больше, чем у животных с полушариями переднего мозга, имеющими борозды и извилины. Так, у бобров поперечник коры несколько больше, чем у человека и медведя, а у дельфина афалины меньше почти в 2 раза. При всём разнообразии вариантов строения неокортекса шестислойность коры млекопитающих не вызывает сомнения (см. рис. III-22; рис. III-23-III-26).
— молекулярный слой (lamina molecularis, lamina zonalis) — самый наружный слой коры, который содержит редкие клеточные тела.
— наружный гранулярный слой (lamina granulans externa) состоит из небольших и плотно расположенных клеточных тел. В моторных областях этот слой может включать небольшие пирамидные клетки, которые распространяют свои волокна на поверхности мозга или в молекулярном слое.
— наружный пирамидальный слой, или слой наружных пирамид (lamina pyramidalis externa). В этом слое преобладают пирамидные нейроны, которые крупнее во внутренних зонах слоя и мельче — в наружных.
— внутренний гранулярный слой (lamina granulans interna). Этот слой состоит из плотно расположенных как пирамидных, так и непирамидных клеток.
— пирамидальный слой (lamina pyramidalis). В этом слое расположены преимущественно клетки округлой формы, которые посылают свои волокна к пирамидным нейронам.
— слой полиморфных клеток (lamina multiformis). В нём находятся веретеновидные, округлые и модифицированные пирамидные клетки.
В неокортексе млекопитающих есть пирамидные клетки, которые отсутствуют в мозге других позвоночных. В моторной коре находятся особенно крупные пирамидные нейроны, носящие название клеток Беца (см. рис. III-24). Эти нейроны посылают свои длинные отростки к мотонейронам спинного мозга и в двигательные ядра черепно-мозговых нервов. Их гигантский размер предопределён масштабами метаболической и функциональной активности.
Неокортекс млекопитающих различается по толщине и выраженности клеточных слоёв. Клеточные слои, образующие кору, могут сливаться вместе или разделяться на несколько подслоёв в зависимости от функциональной принадлежности поля. Даже у относительно близких видов в функционально одинаковых полях неокортекса можно обнаружить цитоархитектонические различия (см. рис. III-26). При этом совершенно не обязательно, чтобы кора высокогирифицированного мозга была организована сложнее, чем мозга лиссэнцефального.
Кора млекопитающих неоднородна даже в пределах одного полушария и имеет различный клеточный состав. Это позволило выделить в коре специфические однотипно организованные центры — цитоархитектонические поля. Каждое морфологическое поле коры соответствует представительству конкретного периферического анализатора, ассоциативной или моторной зоне. Примером может служить неокортекс лиссэнцефального мозга кролика (см. рис. III-24). Он содержит несколько крупных полей, представляющих собой моторные, сенсомоторные, зрительные и ассоциативные центры. Каждое специализированное поле коры имеет цитоархитектонические особенности расположения нейронов, которые могут различаться по форме и размеру. В моторных полях коры есть крупные пирамидные нейроны, отсутствующие в других областях неокортекса (см. рис. III-24).
Читать дальшеИнтервал:
Закладка:



