Константин Саевич - Общая и прикладная экология

- Название:Общая и прикладная экология
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:2014
- Город:Минск
- ISBN:978-985-06-2400-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Константин Саевич - Общая и прикладная экология краткое содержание
Общая и прикладная экология - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Правило Лотки в большей степени справедливо для высших организмов с развитой возрастной структурой популяции и не имеет свойств универсальности, однако в более широком, биосистемном, смысле оно признается универсальным.
При наличии у данного вида организмов половой дифференциации правило стабильности возрастной структуры популяции необходимо дополнять правилом стабильности соотношения полов .
Половая дифференциация (бисексуальность) играет огромную роль в поддержании генетической разнокачественности особей популяции, что, в свою очередь, обеспечивает устойчивость популяции , т. е. ее способность адекватно реагировать на изменения окружающей среды. Половая структура бисексуальной популяции определяется количеством самок и самцов в ней.
Принято выделять следующие соотношения полов в популяции: первичное соотношение полов определяется генетическими механизмами, т. е. равномерностью распределения половых хромосом; вторичное соотношение полов – это соотношение полов на момент рождения; третичное соотношение полов – это соотношение полов среди взрослых (репродуктивных) особей.
Для определения половой структуры популяции существенную роль играет половой индекс.
Половой индекс ( I s ) – это отношение общего числа половозрелых самок ( n ♀ ж) к общей численности популяции ( N ):
I s = n ♀ … / N .
В совокупности эти два рассмотренных правила составляют правило стабильности половозрастной структуры популяции .
Таким образом, плодовитость является константой, определяемой расчетным путем, например умножением среднего числа гнезд, которые способна построить самка птицы за год, на такое же число яиц, которые она может отложить в наиболее благоприятную часть сезона года.
Смертность – величина, противоположная рождаемости, может быть определена как число особей Δ N m , погибших за время Δ t . Так же как и при оценке рождаемости, смертность обычно относят к общему числу особей в популяции N , а промежуток Δ t стараются брать по возможности короче.
М г н о в е н н а я удельная смертность d выражается формулой
d = d N m / N d t .
Размерность мгновенной удельной смертности такая же, как рождаемости. Традиционно величина смертности в экологической и демографической литературе обозначается буквой d (от англ. death-rate ). При этом не следует путать d (смертность) с d (знаком дифференциала в выражениях типа d N /d t ).
Численность и плотность популяции зависит, наряду с рождаемостью, и от смертности.
Смертность популяции – это количество особей, погибших за определенный период. Различают абсолютную (общую) и удельную смертность.
А б с о л ю т н а я (о б щ а я) смертность – это число особей (Δ Nm ), погибших в единицу времени (Δ t ).
У д е л ь н а я смертность ( d ) выражается отношением абсолютной смертности к численности популяции:
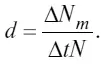
Абсолютная и удельная смертность характеризуют скорость убывания численности популяции вследствие гибели особей от хищников, болезней, старости и т. д.
Экологическая ( реализованная ) смертность – гибель особей в данных условиях среды. Эта величина не постоянная, она изменяется в зависимости от условий среды и состояния популяции. Полная картина смертности описывается статистическими таблицами выживаемости.
Смертность является величиной положительной или равной нулю, но последнее бывает крайне редко и только в течение короткого времени.
Различают три типа смертности:
• первый – характеризуется одинаковой смертностью во всех возрастах. Выражается экспоненциальной кривой (убывающей геометрической прогрессии). Данный тип смертности встречается редко и только у популяций, которые постоянно находятся в оптимальных условиях (человек, некоторые млекопитающие, дрозофилы и другие лабораторные животные и растения);
• второй – характеризуется повышенной гибелью особей на ранних стадиях развития и свойственен большинству растений и животных. Максимальная гибель животных происходит в личиночной фазе или в молодом возрасте, у многих растений – в стадии произрастания семян и всходов. У насекомых до взрослых особей доживает 0,3–0,5 % отложенных яиц, у многих рыб – 1–2 % количества выметанной икры;
• третий – отличается повышенной гибелью взрослых, в первую очередь старых особей. Распространен у насекомых, личинки которых обитают в почве, воде, древесине, а также в других местах с благоприятными условиями защиты, тогда как взрослые особи подвергаются постоянным воздействиям неблагоприятных факторов среды.
В связи с этим в экологии широко распространено графическое построение кривых выживания (рис. 1.4).
Располагая по оси абсцисс продолжительность жизни в процентах от общей продолжительности жизни, можно сравнивать кривые выживания организмов, продолжительность жизни которых имеет значительные различия. На основании таких кривых определяют периоды, в течение которых тот или иной вид особенно уязвим.
Если действие причин, вследствие которых изменяются рождаемость или смертность, приходится на наиболее уязвимую фазу, то их влияние на последующее развитие (рост численности, плотность, плодовитость и т. д.) будет максимальным. Это позволяет, например, регулировать численность насекомых-вредителей или грызунов в зернохранилищах.
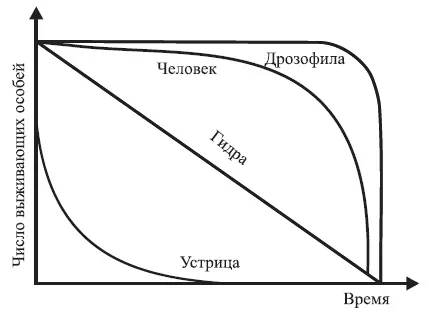
Рис. 1.4. Различные типы кривых выживания (по Р. Дажо, 1975)
Смертность подвержена более резким колебаниям и больше зависит от факторов окружающей среды, чем рождаемость, она играет главную роль в регулировании численности популяции.
Численность популяции является видоспецифическим признаком.
В любой природной системе поддерживается та численность особей в популяциях обитающих здесь животных и растений, которая в наибольшей степени отвечает интересам воспроизводства популяций.
Основное уравнение динамики численности популяции выглядит следующим образом:
r = b – d ,
где r – скорость наблюдаемого изменения численности, или коэффициент прироста изолированной популяции; b – коэффициент рождаемости; d – коэффициент смертности.
В соответствии с этим уравнением рост популяции описывается экспоненциальной кривой. Это говорит о том, что оно справедливо для идеальной популяции, имеющей неограниченные пищевые ресурсы, не подвергающейся никаким внешним воздействиям среды и распространяющейся на неограниченной территории.
Читать дальшеИнтервал:
Закладка:







