Несса Кэри - Мусорная ДНК. Путешествие в темную материю генома

- Название:Мусорная ДНК. Путешествие в темную материю генома
- Автор:
- Жанр:
- Издательство:Лаборатория знаний
- Год:2016
- Город:Москва
- ISBN:978-5-906828-62-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Несса Кэри - Мусорная ДНК. Путешествие в темную материю генома краткое содержание
Расшифровав генетический код, ученые обнаружили, что лишь 2% ДНК несут информацию о белках. А для чего же тогда нужны оставшиеся 98%? Поначалу генетики решили, что это мусор, хлам. Однако совсем недавно стало ясно — все гораздо сложнее, и именно эти «мусорные» области ДНК определяют сложность человеческого организма, его возможные болезни и даже — скорость старения! Здесь — ключи к пониманию эволюции и сущности самой жизни.
Сегодня множество ученых в самых разных лабораториях мира пытаются проникнуть в тайны «мусорной» ДНК, этой темной материи нашего генома. Об их последних результатах — в увлекательной книге английского генетика Нессы Кэри.
Мусорная ДНК. Путешествие в темную материю генома - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Поскольку плюрипотентное состояние ЭС-клеток во многом определяется высокими уровнями экспрессии главных регуляторов, неудивительно, что сами главные регуляторы контролируются суперэнхансерами. Это создает систему с положительной обратной связью (см. рис. 12.2).
Циклы с положительной обратной связью сравнительно редко встречаются в биологии — главным образом из-за того, что в том случае, если они начнут вести себя как-то не так, их трудно будет снова обуздать. По счастью, кодирующие белок гены, регулируемые суперэнхансерами, весьма чувствительны к малейшим изменениям связывания главных регуляторов и к целому ряду других факторов. А значит, не исключено, что даже небольшого изменения в балансе этих факторов может оказаться достаточно для прерывания этого цикла с положительной обратной связью, что позволит клеткам не оставаться плюрипотентными, а начать дифференциацию. В конце концов, обычно не требуется больших усилий, чтобы столкнуть пружину «слинки» с лестницы.
Сообщалось также о наличии суперэнхансеров в клетках злокачественных опухолей. Вероятно, там они взаимодействуют с генами, оказывающими ключевое воздействие на размножение клеток и развитие раковых процессов 16. Один из генов, регулируемых таким суперэнхансером, порождает лимфому Бёркитта (мы уже упоминали этот ген в данной главе). В кое-каких нормальных специализированных клетках тоже имеются суперэнхансеры. Они связываются с белками, специфическими для клеток данного типа и определяющими особенности клетки.
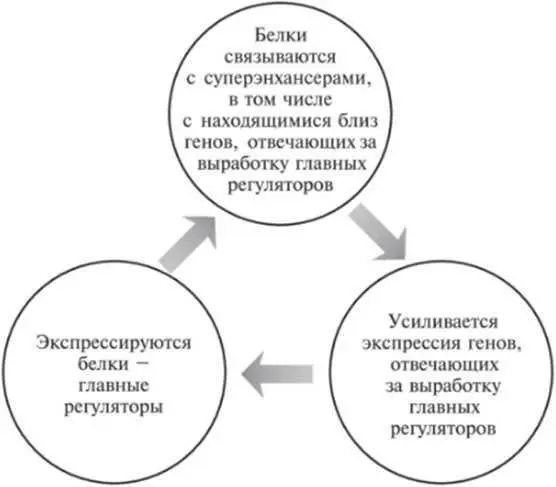
Рис. 12.2.Цикл с положительной обратной связью, поддерживающий высокий уровень экспрессии генов, которые отвечают за выработку главных регуляторов.
Преодолевая расстояния
В большинстве событий, которые мы описывали в этой главе, участвуют энхансеры, которые находятся сравнительно близко от таргетируемых ими генов: обычно в пределах 50 тысяч нуклеотидных оснований. Довольно легко представить себе, как это происходит: длинная некодирующая РНК и комплекс-медиатор действуют как своего рода якорь для фермента, копирующего ДНК в информационную РНК. Однако во многих ситуациях энхансер и регулируемый им ген, кодирующий белок, на хромосоме отстоят друг от друга довольно далеко: их может разделять до нескольких миллионов пар нуклеотидных оснований. Такая же разница, как между попыткой передать солонку кому-то на противоположном конце стола и попыткой передать ее кому-то, кто располагается на другом конце футбольного поля. Нелегко представить себе, каким образом может осуществляться такого рода дистанционное взаимодействие между геном и энхансером. Размеров длинной некодирующей РНК и комплекса-медиатора не хватит, чтобы перекрыть такое огромное расстояние.
Чтобы разобраться в этом процессе, придется усложнить обычный образ генома. Как правило, весьма полезно и удобно описывать ДНК как веревочную лестницу или железнодорожные пути, ибо это помогает вообразить две ее нити и то, как они удерживаются вместе посредством пар нуклеотидных оснований. Однако проблема в том, что при этом мы думаем о ДНК как о чем-то линейном, к тому же достаточно жестком, поскольку подсознательно мы сравниваем ее с твердыми предметами из более привычной нам среды.
Но мы уже знаем, что ДНК — не жесткая молекула, ведь ее можно очень сильно сплющивать, сминать, комкать, чтобы она уместилась в ядре. Так что давайте углубим наши исследования. Если все-таки принять двунитевую природу ДНК как данность (чтобы не усложнять картину), можно представить себе фрагмент нашего генома как длинную макаронину — возможно, самый длинный в мире кусок лапши тальятелле. В двух местах она отмечена пищевым красителем: эти места изображают энхансер и ген, кодирующий белок. Глядя на рис. 12.3, мы видим два возможных сценария. Пока макароны не сварили, они сохраняют жесткость, и энхансер находится вдали от гена. Но после варки макароны становятся гибкими. Теперь они могут сгибаться во всевозможных направлениях, в результате чего окрашенные участки, изображающие энхансер и ген, могут сближаться.
Некоторые части наших хромосом в определенных клетках подавляются и выключаются почти навсегда — чтобы отключить гены, которые никогда не понадобится экспрессировать в тканях данного типа. Скажем, клеткам нашей кожи незачем экспрессировать белки, которые используются кровью для переноса кислорода. В клетках кожи соответствующие области генома совершенно недоступны: они плотно свернуты и напоминают чересчур сжатую пружину. Однако в этих же клетках есть громадные области, не находящиеся в таком сверхсжатом состоянии. Гены в них вполне доступны и могут включаться. ДНК в таких зонах напоминает самую длинную вареную макаронину в мире, целиком заполняющую кастрюлю. Макаронина сгибается и извивается в кипящей воде, образуя всевозможные петли и дуги.
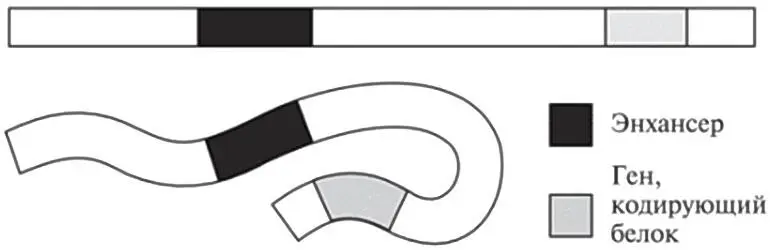
Рис. 12.3.Упрощенная схема показывает, как складывание гибкой молекулы ДНК может сблизить два отдаленных ее участка — скажем, энхансер и ген, кодирующий белок.
В таких случаях ген, кодирующий белок, и расположенный вдали от него энхансер могут сильно сближаться друг с другом. Затем длинная некодирующая РНК и комплекс-медиатор удерживают две петли вместе, обеспечивая усиление экспрессии гена. В этом медиатору должен помогать еще один комплекс [39] Этот белковый комплекс называется когезином.
. Этот дополнительный комплекс требуется также для разделения хромосом, прошедших дупликацию в процессе деления клетки, так что он хорошо оснащен для работы с масштабными перемещениями ДНК. Мутации в некоторых генах, кодирующих белки этого дополнительного комплекса, вызывают две болезни развития — синдром Робертса и синдром Корнелии де Ланге 17. Их проявления могут быть довольно разными и, вероятно, определяются тем, какой именно ген мутирует и какова эта мутация. Обычно дети с этими заболеваниями рождаются слишком маленькими и в дальнейшем сохраняют это отставание в росте. Они испытывают трудности с обучением. Зачастую у них деформированы конечности 18.
Подобный «петлевой» механизм распространен довольно широко. Возможно, он применим не только к энхансерам. Не исключено, что с его помощью и другие регуляторные элементы сближаются с теми или иными генами. В рамках исследования трех клеточных типов, затронувшего лишь 1% человеческого генома, удалось выявить более 1000 таких дистанционных взаимодействий в каждой линии клеток. Эти сложные взаимодействия чаше всего связывали области, разделенные примерно 120 тысячами пар нуклеотидных оснований. Среди более чем 90% таких петель ближайший ген просто игнорировался. Представьте, что вам понадобилось одолжить немного сахара, и вы отправляетесь не к ближайшему соседу, а к кому-нибудь, кто живет в полумиле от вас.
Читать дальшеИнтервал:
Закладка:






![Надежда Алданен - Темные Материи [Dark Matters] II Заговор](/books/1158689/nadezhda-aldanen-temnye-materii-dark-matters-ii-z.webp)


