Михаил Супотницкий - Микроорганизмы, токсины и эпидемии

- Название:Микроорганизмы, токсины и эпидемии
- Автор:
- Жанр:
- Издательство:Вузовская книга
- Год:2006
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Супотницкий - Микроорганизмы, токсины и эпидемии краткое содержание
Кроме того, в монографии, впервые для отечественной литературы, обстоятельно освещены эпидемиология биологического террористического акта и методология выявления такого преступления. Книга хорошо иллюстрирована, содержит приложения и рассчитана на широкий круг специалистов.
Микроорганизмы, токсины и эпидемии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
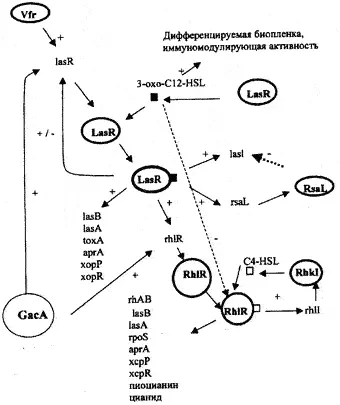
Рис. 9. Схематическое изображение регуляции генов патогенности Pseudomonas aeruginosa. Система сигналов las, передаваемых от «клетки к клетке», в иерархическом каскаде контролируется посредством сигнальной системы белка rhi Комплекс LasR/-3-oxo-C12-HSL активирует транскрипцию rhlR. 3-охо-12HSL блокирует активацию RhlR посредством C4-HSL. Сама las-система контролируется позитивно — посредством Vfr и GacA, и негативно — посредством RsaL. 3-oxo-C12-HSL требуется для дифференциации биопленки и обладает иммуномодуляторной активностью. Обе сигнальные системы регулируют экспрессию различных генов (lasB — LasB-эластаза; lasA — LasA-эластаза; toxA — экзотоксин А; аргА — щелочная протеаза хсрР и xcpR — гены хср-секреторного пути; rhlАВ — рамнозилтрансфераза, требуемая для продукции рамноллипида; rpoS — сигма-фактор стационарной фазы [Van Delden С, Iglewski В., 1998]
Shigella flexneri — возбудитель дизентерии у людей. Посредством эндоцитоза, через лимфатические фолликулы пейферовых бляшек тонкого кишечника, шигеллы проникают в слизистую оболочку ободочной кишки (рис. 10). Продуктами генов, которые инициируют процесс инвазии, являются три белка: IpaB (плазмидный антиген инвазии — 62 кд), IpaD (38 кд), IpaC (42 кд). В lamina prdpria они инфицируют макрофаги и вызывают запрограммированную гибель клеток. Цитокины высвобождаются и индуцируют воспаление. Вследствие притока полиморфно-ядерных фагоцитов разрушается базальная мембрана и разрывается эпителий. Клетки, теперь уже лишенные щеточной каемки, становятся уязвимыми и не защищенными от индуцируемого плазмидой проникновения шигелл. Детерминантой апоптической гибели макрофагов является белок IpaB. В течение короткого времени после проникновения шигелл в клетки, вакуоли лизируются под действием IpaB. Высвободившись, бактерии быстро размножаются внутри цитоплазмы. Шигеллы неподвижны и лишены жгутиков, но они перемещаются внутри клетки и распространяются от одной клетки к другой. Это обусловлено образованием у них «хвоста» из полимеризованного актина хозяина под влиянием гена ics (гена внутриклеточного распространения), который кодирует 120 кд белок. Перенос шигелл осуществляется через выступы на поверхности одной клетки, которые вставлены в инвагинации соседней клетки. Затем части, содержащие бактерии, отсекаются. Двойные мембраны лизируются продуктами плазмидного гена icsB, высвобождая бактерии для дальнейших циклов роста и переноса [Smith H., 1995; Finlay В Falkow S., 1997; Goosney D. et al., 1999].

Рис. 10. Механизм инвазии Shigella flexneri. A. Иммунофлюоресцентная фотография, показывающая Shigella (темноевключение), проникающую через цитоплазму посредством полимеризованного актина (светлые нити). B. Перестановки цитоскелета, обусловленные инвазией Shigella. Наружный мембранный белок IcsA используется шигеллой для продвижения вклетку хозяина. IcsA непосредственно соединяется с двумя белками клетке хозяина — винкулином и белком нейрального синдрома Вискота-Аль-Дрехта (N-WASP). Шигелла расщепляет винкулин, в результате образуется 90 кд фрагмент, который присоединяется к IcsA и к вазодилататорстимулирующему белку (VASP). VASP рекрутирует белки цитоскелета клетки хозяина (актин и профилин) к бактериальной поверхности и формирует актиновый «мотор» для продвижения шигеллы
Salmonella typhimurium — возбудитель энтероинфекции людей (рис. 11). Для проникновения возбудителя в нефагоцитирующий эпителий необходимы несколько хромосомных генов (inv/spa), кластированных на «острове патогенности», названном SPI1 (Salmonella pathogenicity island 1; более подробно см. «острова патогенности» и «системы секреции бактериальной клетки»). Подобно ЕРЕС, АРМ кодирует третий тип секреторной системы, активируемой посредством межклеточного контакта. Это позволяет экспортировать и клетку хозяина детерминанты вирулентности, необходимые для бактериальной инвазии [Goosney D. et al., 1999].
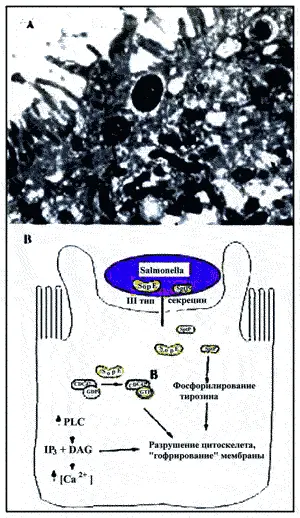
Рис. 11. Механизм инвазии Salmonella typhimurium. A. Трансмиссионная электронная микрофотография индуцированного Salmonella «рифления» мембраны поляризованных Сасо-2 эпителиальных клеток. B. Инвазия Salmonella в эпителиальные клетки хозяина. Salmonella секретирует белки вирулентности, включающие SopE и SptP с помощью секреторной системы III типа. SopE функционирует как фактор обмена гуанидина для небольших СТР(гуанидинтрифосфат) — связывающих белков, вероятно, вызывая обмен GDP на GTP белка CDC42, члена семейства Rho. SptR является фосфатазой, необходимой для инвазии. Предполагается, что она разрушает цитоскелет. Инвазия также стимулирует активность фосфолипазы С (PLC1, приводящей к истечению из клетки и иозитолтрифосфата (IP3) и Са 2+. Последний, в свою очередь, «обратным ходом» может быть вовлечен в перестройку цитоскелета, что приводит к «рифлению» мембраны эукариотической клетки и интернализации Salmonella [Goosney D. et al., 1999]
Yersinia. Наиболее изучены системы инвазии у Yersinia enterocolitica и у Yersinia pseudotuberculosis. Первоначально они проникают в организм через посредничество М-клеток пейеровых бляшек подвздошной кишки. Дальнейшая диссеминация происходит благодаря выживанию внутри макрофагов, которые мигрируют через лимфатическую систему. Оба микроорганизма обладают хромосомными генами, кодирующими наружный мембранный белок инвазии, способствующий их адгезии и проникновению в нефагоцитирующие клетки. Показано, что инвазин эффективно присоединяется к белкам семейства (бета1-интегринов. После тесного связывания с интегринами, инвазин индуцирует проникновение бактерии во внутрь клетки с помощью механизма, подобного «застежке молнии» («zipper-like» mechanism — «зип-лайк» механизм). Он заключается в «расстегивании» мембраны вокруг бактерии в момент ее проникновения в клетку хозяина. Энтеропатогенные Yersinia обладают еще двумя инвазинами: Ail и YadA. Белок Ail способствует более эффективной адгезии к эпителиальным клеткам, но не участвует в их инвазии. Зато он способствует устойчивости Yersinia к действию сыворотки (такие белки широко распространены среди других инвазивных бактерий). YadA — это белок, закодированный на плазмиде. Подобно инвазину он связывается с (бета1-интегринами клеток хозяина. Присоединение Yersinia к бета1-интегрину клетки хозяина запускает механизм контакт-зависимой секреции плазмидных факторов вирулентности и последующей транслокации в ее цитоплазму отдельных бактериальных белков [Smith H., 1995; Finlay В, Falkow S., 1997].
Патогены, связывающие молекулы хозяина для осуществления инвазии. Механизмы инвазии отдельных патогенов не поддаются логике. Легионеллы и микобактерии связывают фрагменты комплемента СЗb и СЗbi, которые облегчают их проникновение в фагоцитирующие клетки, и тем самым, уменьшают для них риск подвергнуться воздействию окислительных радикалов [Schlesinger L.S., Haas R., 1994]. Другие микроорганизмы связывают фибронектин, который затем функционирует как мостик между бактерией и фибронектиновым рецептором клетки хозяина, способствуя их инвазии. Например, Mycobacterium leprae продуцирует фибронектинсвязывающий белок, который способствует ее проникновению в эпителий и шванновские клеточные линии [Schorey А.В. et al., 1995]. Интересно, что механизмы, используемые для инвазии в нефагоцитирующие клетки функционируют и в отношении фагоцитирующих клеток. Например, мутант S. typhimurium, утративший способность к инвазии эпителиальных клеток, одновременно значительно снижает свою способность проникать в фагоцитирующие клетки [Gahring L.C., 1990]. Это возможно потому, что использование бактериями путей активной инвазии, помогает им избежать антибактериальной активности фагосом, куда они неизбежно попадают при традиционном фагоцитозе [Finlay В., Falkow S., 1997].
Читать дальшеИнтервал:
Закладка:





![Михаил Супотницкий - Очерки истории чумы. Книга II. Чума бактериологического периода [без иллюстраций]](/books/1084186/mihail-supotnickij-ocherki-istorii-chumy-kniga-ii.webp)
![Михаил Супотницкий - Очерки истории чумы. Книга I. Чума добактериологического периода [без иллюстраций]](/books/1084187/mihail-supotnickij-ocherki-istorii-chumy-kniga-i-ch.webp)


