Михаил Супотницкий - Микроорганизмы, токсины и эпидемии

- Название:Микроорганизмы, токсины и эпидемии
- Автор:
- Жанр:
- Издательство:Вузовская книга
- Год:2006
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Супотницкий - Микроорганизмы, токсины и эпидемии краткое содержание
Кроме того, в монографии, впервые для отечественной литературы, обстоятельно освещены эпидемиология биологического террористического акта и методология выявления такого преступления. Книга хорошо иллюстрирована, содержит приложения и рассчитана на широкий круг специалистов.
Микроорганизмы, токсины и эпидемии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Нестабильность, как и наоборот, стабильность «островов», видимо не случайность в проявлении бактерией патогенности. Эти свойства способны создать бактерии дополнительные адаптивные преимущества. Высокая вирулентность может оказаться невыгодной бактерии на определенной стадии инфекционного процесса. Нестабильность же «островов патогенности» будет способствовать координированному снижению вирулентности всей популяцией возбудителя инфекции. Делеция «островов патогенности» может усилить экспрессию других, рядом расположенных, генов [Ritter A., 1995].
С другой стороны, отдельные факторы вирулентности являются адаптивными для бактерий. По этой причине они должны кодироваться на стабильных «островах патогенности». «Чужеродность» «островов» придает даже большую стабильность факторам вирулентности. Чужеродная ДНК, интегрировавшаяся с хромосомой, не вовлекается в рекомбинацию с ДНК близкородственных микроорганизмов, поэтому она может длительно поддерживаться в бактериальных популяциях [Falkow S., 1996]. Видимо применительно к каждому патогенному микроорганизму «стабильность» и «нестабильность» «островов» закрепляется естественным отбором.
В настоящее время нет полной ясности, каким образом «острова патогенности» приобретаются бактериями, но есть косвенные признаки того, что этот процесс идет непрерывно. Новый эпидемический штамм Vibrio cholerae О139 появился в результате приобретения «острова патогенности». Хотя известно, что он возник из штамма того же серотипа (О1), который является причиной происходящей в настоящее время пандемии холеры (О1 El Tor), V. cholerae О139 содержит дополнительный участок ДНК, который замещает часть О-антигенного кластера О1-штамма. Вставленная ДНК содержит открытую рамку считывания, гомологичную протеину, вовлеченному в синтез капсулы и О-антигена — двух факторов, по которым можно провести различия между О139 и О1 Е1 Тог. Тем самым повышается способность возбудителя холеры к инвазии [Mecsas J. Strauss E.J., 1996].
Бактерии являются мощными раздражителями иммунной системы, поэтому они могут использовать только первую стратегию паразитизма и в иммунокомпетентном хозяине лишь кратковременно имитировать вторую стратегию (L-формы). Бактерии обладают избыточными и альтернативными механизмами адгезии и колонизации, что позволяет им проявлять большую гибкость в выборе новых хозяев. Адгезия бактерий не является простым механическим взаимодействием их лиганд-структур с рецепторами на поверхности клеток-мишеней хозяина, имеющими другое предназначение. Взаимодействие патогена с клеткой хозяина приводит к активации сигнальных систем клеток-мишеней непосредственно каким-то бактериальным компонентом, либо через стимуляцию активационных факторов хозяина. Для противодействия иммунной системе бактерии используют как механизмы уклонения (экранирование клеточной стенки бактерий, антигенная мимикрия, образование L-форм бактерий и др.), так и разрушения (продуцирование секретируемых факторов, инактивирующих защиту хозяина; продуцирование «суперантигенов» и др.). Гены, кодирующие факторы патогенности бактерий и системы их секреции, вопреки распространенному еще недавно мнению об их преимущественно плазмидной локализации, в большинстве своем кластированны на участках хромосомной ДНК, называемых «островами патогенности». Эти участки активно участвуют в генетическом обмене между бактериями по пока еще неизвестному механизму. Патогенность «новых» для человека бактерий может объясняться тем, что они уже были преадаптированы к такой встрече в занимаемых ими ранее экологических нишах.
Глава 1.4. Патогенность микоплазм
Происхождение микоплазм. Патогенность микоплазм. Болезни, вызываемые микоплазмами.
Микоплазмы — это уникальные прокариотические организмы, имеющие лишь одну липопротеиновую мембрану, которая выполняет функции и клеточной стенки, и цитоплазматической мембраны. Они входят в класс Mollicutes, объединяющий также и ахолеплазмы, спироплазмы, уреаплазмы, анаэроплазмы [Прозоровский СВ. и др., 1995]. По форме и структурной организации молликуты очень близки к L-формам бактерий [Бухарин О.В., Усвяцов Б.Я., 1996]. Но в отличие от них, геном молликут крайне упрощен и экономен, что не предполагает возможность реверсии клеточной стенки. Микоплазмы являются самыми маленькими самореплицирующимися формами жизни и способны вызывать инфекционный процесс, используя вторую стратегию паразитизма. Болезни, вызываемые микоплазмами, называются микоплазмозами.
Происхождение микоплазм. Филогенетические связи класса Mollicutes и пути их эволюции обсуждаются специалистами в основном с двух точек зрения. Согласно одной из них микоплазмы являются выжившей ветвью примитивных организмов, из которых впоследствии произошли прокариоты и эукариоты. Они появились, как продукт прогрессивной эволюции еще до образования присущей бактериям клеточной стенки. Другая точка зрения состоит в том, что микоплазмы являются регрессивной ветвью эволюции некоторых грамположительных бактерий и (или) клостридий. Второе предположение находит экспериментальные подтверждения и рассматривается в двух возможных вариантах. Все микоплазмы происходят либо из одного предка, общего с грамположительными бактериями, либо от разных бактерий [Прозоровский СВ. и др., 1995].
Патогенность микоплазм. Так же как и бактерии, микоплазмы, что бы быть патогенными, должны инфицировать поверхности клеток хозяина. Поэтому многие патогенные микоплазмы имеют форму нитей или колбочек и на их поверхности, при электронной микроскопии, видны кончики выступающих полярных органелл, участвующих в процессе адгезии к клетке хозяина [Kirchhoff H. et al., 1984]. Эти концевые структуры имеют сложное строение. Они сплетены из интерактивных белков адгезинов и белков, участвующих в адгезии. Интересной особенностью адгезинов микоплазм является генетическое регулирование их антигенной структуры (рис. 14 и 15).
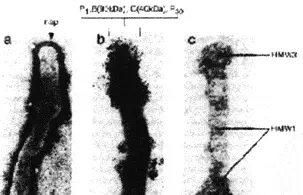
Рис. 14. Концевая структура органеллы, участвующей в адгезии микоплазмы. Трансмиссионная электронная микрофотография М. pneumoniae: a) скрученная структура концевой части органеллы с пар (участок адгезии); b) кластированые цитоадгезиновые белки (Р1, В, С, Р30) в иммунолабильном участке концевой части органеллы (окрашивание ферритином и коллоидальным золотом); с) резистентная к Тритону Х-100 цитоскелетоподобная структура с отчетливыми пузырьками и параллельными нитями [Baseman J., Tully J., 1997]
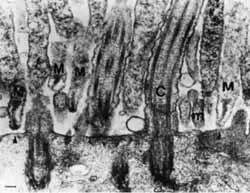
Рис. 15. Адгезия микоплазм к трахеальному эпителию хомячка.
Читать дальшеИнтервал:
Закладка:





![Михаил Супотницкий - Очерки истории чумы. Книга II. Чума бактериологического периода [без иллюстраций]](/books/1084186/mihail-supotnickij-ocherki-istorii-chumy-kniga-ii.webp)
![Михаил Супотницкий - Очерки истории чумы. Книга I. Чума добактериологического периода [без иллюстраций]](/books/1084187/mihail-supotnickij-ocherki-istorii-chumy-kniga-i-ch.webp)


