К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Моя реакция на изобилие экспериментальных данных, полученных при исследовании энграммы в 1960-е гг., двойственна. Во-первых, можно прийти к выводу, что существует не один механизм памяти, не единая молекула памяти. Пластичность нейрона многообразна, и память не является единым процессом. Воображение, узнавание, припоминание предполагают использование некоторых или всех основных форм пластичности нервной ткани и различных способов сохранения и воспроизведения следа. Поэтому первый вывод, который следует сделать, состоит в том, что память, как она обычно представляется, не единый механизм, лежащий в основе процессов, которые позволяют организму устанавливать связи между отдельными звеньями опыта.
Во-вторых, простая модель следов, предполагающая постоянную модификацию мозговой ткани на нейронном уровне, может быть сформулирована уже сейчас. Такая модель должна обеспечить не только возможность сохранения следов, но и доступность считывания зафиксированного изменения. Наконец, такая модель должна быть основана на уже имеющихся данных и отвечать здравому биологическому смыслу. В настоящее время наиболее вероятным источником такой модели может служить аналогия с процессом, происходящим во время эмбрионального развития организма (рис. II-13).

Рис. II-13. Связь в виде треугольника, сформированная между тремя спинальными ганглиями эмбриона in vitro. Рисунок иллюстрирует тенденцию нервной ткани формировать узоры нервных волокон (Weiss, 1967).
Такие структуры, как глаз, формируются только тогда, когда ткань, из которой они развиваются, соответствующим образом стимулируется. Большая часть эмбриональной ткани эквипотенциальна, то есть ДНК во всех клетках тела, по существу, одна и та же. Это означает, что потенциал первоначально подавлен, находится в состоянии «репрессии». «Дерепрес-сия», или освобождение потенциальных возможностей, происходит тогда, когда появляются соответствующие условия. Определение эмбриологами того, каковы эти условия, заняло почти целое столетие. В настоящее время хорошо известно, что РНК и некоторые эндокринные секреты являются индукторами такой «дерепрессии». Маловероятно, что эти химические соединения несут в себе всю информацию, необходимую для индукции; большая доля специфики процесса должна быть отнесена за счет субстрата. Например, РНК, выделенная из печени теленка, может быть использована как индуктор для хрусталика глаза крысы. Отношения между индуктором и субстратом, по-видимому, следующие.
1. Индукторы извлекают и реализуют потенциальную генетическую информацию организма
2. Индукторы в известной мере специфичны в отношении характера извлекаемой генетической информации, но они неспецифичны относительно вида особи или ткани
3. Индукторы определяют общую схему индуцированного свойства, специализация деталей возникает в результате деятельности субстрата
4. Индукторы непосредственно не являются толчком для развития, они составляют особый класс стимулов
5. Чтобы быть эффективными, индукторы должны взаимодействовать с субстратами Однако, чтобы вызвать эффект, недостаточно простого контакта – ткань должна быть готова правильно реагировать
6 Индукция обычно развивается в результате действия в двух направлениях благодаря химическому взаимодействию между индуктором и субстратом.
Данные о роли РНК в сохранении следов памяти в настоящее лремя могут быть объяснены при помощи модели, основанной на этом эмбриональном процессе индукции. Модель предполагает, что возбуждение нервных структур сопровождается образованием РНК. Эта нейронная РНК вызывает изменения в окружающей «лигодендроглии, в результате чего начинается химический процесс между нейроном и глией, характеризующийся реципрокными отношениями изменений нейронной и глиальной концентрации РНК (и множества метаболитов). Происходит изменение в функциональном взаимодействии между глией и нейроном. На первом этапе может возникнуть соответствующее изменение концентрации РНК в глиальной клетке, которое затем в течение длительного времени будет вызывать изменение в структурах липидов, протеинов и липопротеинов во всех больших молекулах, составляющих мембраны, через которые осуществляется взаимодействие нейрона с глией. Такие макромолекулярные изменения могут влиять на легкость освобождения и разрушения нейронных медиаторов. Эти структурные изменения обратимы и могут стираться или быть вытеснены другими изменениями. Однако, если при неоднократном повторении одного и того же типа воздействия' изменения в молекулярной структуре будут продолжаться достаточно долго, они вызовут эффективные изменения мембранной проницаемости, которые в свою очередь способствуют появлению в возбужденном нейроне большого количества РНК, метаболитов и медиаторов, оказывающих влияние на окружающую его глию в том месте, где происходит деление глиальной клетки. Конус роста нейрона, освободившийся от инкапсулированной глии, свободно воздействует на недавно сформировавшиеся дочерние клетки в образует новые связи с нейронами. Таким образом, клеточное деление олигодендроглии направляет конусы роста нейронов центральной нервной системы во многом так же, как на периферии их рост направляют соответствующие шванновские клетки (рис. II-14).
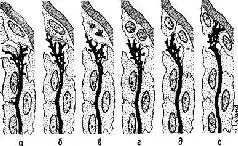
Рис. II-14 Шесть фаз гипотетического процесса индукции
Любое изменение глиально-нейронной организации следует рассматривать как изменение в микроструктуре активности медленных потенциалов соединений. Как мы уже говорили, такие изменения могут быть временными либо вести к возникновению более устойчивых изменений в нервной системе, связанных с ростом нейронов. Имеется много данных, свидетельствующих о том,, что для фиксации в мозгу следов памяти – энграмм – требуется время. Эти данные вполне согласуются с индукционной моделью хранения следов памяти, потому что они указывают на возможность выделения по крайней мере двух – если не больше – процессов в зависимости от времени их появления.
После достаточно сильной травмы головы человек не способен воспроизвести в памяти события, происходившие в период, непосредственно предшествовавший травме. Длительность этого периода зависит от тяжести травмы. Этот феномен, названный ретроградной амнезией, может быть использован для изучения фиксации следов, то есть для изучения процессов консолидации.
Читать дальшеИнтервал:
Закладка:








