К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Конечно, здесь отмечено не все, что свидетельствует о необходимости регистрации волновых форм. Структурные изменения в макромолекулах обладают способностью к восстановлению. Вероятно, более устойчивая запись требует таких механизмов, как настройка «усредняющих циркуляции» в кортикальных колонках и рост, вызванный изменениями в мембранной проницаемости вследствие и в зависимости от этих макромолекулярных изменений. «Фильтр», или «экран», топографических узоров состоит нетолько из решетки макромолекул мембраны, образующих синаптически-дендритную сеть, но также из облегчения всех тенденций формировать образ и создавать выходные динамические структуры нервных импульсов.
Одна группа данных в пользу голографической гипотезы идет от исследований развития зрительного восприятия у новорожденных. Вопреки широко распространенным представлениям (например, Хебб) константность величины и формы не возникает исключительно из комбинаций следов прошлых и, по-видимому, более элементарных восприятий (например, восприятие длины линий, углов). Предположение Хебба основывалось на результатах экспериментов и клинических наблюдений, когда испытуемые находились в условиях сенсорной депривации. Он считал, что для соответствующего развития рецепторной функции необходим опыт. Однако электрофизиологические эксперименты (воспроизведенные на рисунке VII-12) показывают, что сетчаточные и нейронные механизмы, необходимые для детекции признаков, уже хороша развиты у млекопитающих (кошек) к моменту рождения. Поэтому эффект, наблюдающийся при ограничении сенсорных воздействий, по-видимому, является следствием либо атрофии, возникающей в результате бездействия синапсов, либо активного подавления сенсорного механизма посредством вызванного ненормального функционирования. Как уже отмечалось, у человека возникает аналогичная ситуация. Бауер (1966) проводил эксперименты с шестинедельными младенцами, и эти эксперименты показали, что у них уже наблюдается константность величины ж в значительной степени константность формы. В этих экспериментах предметы помещались таким образом, что их действительная величина, расстояние до них и сетчаточный образ (который вычисляется исходя из действительной величины рассматриваемой фигуры) могли образовывать пары в различных сочетаниях. Использовалось полное и незаконченное изображение фигур-Плоское двухмерное изображение предметов заменяло трехмерные предметы. При обучении младенца различению экспериментатор в качестве подкрепления использовал игру в прятки. Затем производилась проверка, которая состояла в выявлении у младенца заученного ответа при замене одного стимула пары другим. Результаты экспериментов показали, что константность возникает у младенца только тогда, когда он располагает сведениями о расстоянии или когда один элемент фигуры перемещается,, тогда как остальные остаются на прежнем месте. Таким образом, даже у младенцев зрительная система представляет собой врожденную организацию, регистрирующую «такой вид информации, который порождается двигательным и бинокулярным параллаксом». Какой же тип встроенного механизма мог бы регулировать параллакс лучше, чем голографический процесс, который сам конструируется посредством параллактического процесса (эффекта интерференции)?
Прямым доказательством того, что в формировании образов участвует параллактический кортикальный механизм, подобный голограмме Фурье, являются результаты экспериментов, проверенных Кемпбеллом и его сотрудниками (Blakemore and Campbell, 1969; Campbell et al. 1968; Campbell, Cooper and Enroth-Cugell, 1969; Campbell and Robson, 1968). Эти исследования показали, что кора головного мозга может настраиваться на восприятие пространственной информации разного диапазона. Эксперименты -были проведены на кошках и на людях. Предъявлялись решетки с различной контрастностью и изучалось влияние их вращения на зрительные вызванные потенциалы (у человека) и на динамические структуры разрядов, возникающих в нейронах зрительной коры (у кошки). Нейроны отвечали на ограниченную полосу пространственной частоты (в четыре октавы), а длительное рассматривание вызывало подавление контрастной чувствительности за пределами соседних частот (рис. VIII-12). Взаимоотношение между этими экспериментальными результатами и голограммой Фурье лучше всего сформулировано самими авторами.
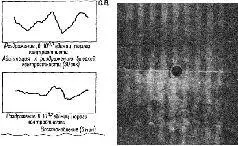
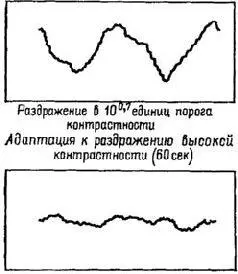
Рис. VIII-12. Эффект пространственной адаптации на вызванные потенциалы у человека. Синусоидально-волновой узор решетки (12 кол/градус) сдвигался вокруг своей оси на 180° со скоростью 8 раз в сек. Вызванный потенциал зрительной коры суммировался 200 раз на специальном аппарате для получения записей, показанных в рамках слева. Для всех записей стимул был одинаковым. Каждая кривая показывает потенциал для двух фазовых сдвигов пространственной решетки. Первая запись получена при рассмотрении решетки низкой контрастности (верхняя часть справа расположенной панели), составляющей 10 0 ' 7 порогов контрастности для этой пространственной частоты. Затем испытуемый в течение 30 сек рассматривал решетку высокой контрастности (на 1,5 единицы выше порога), после чего немедленно производились записи вызванного потенциала в ответ на повторное предъявление решетки с тем же низкич контрастом. Эта вторая кривая имеет гораздо меньшую амплитуду, чем первая. После 5-минутного периода восстановления предъявление решетки низкой контрастности вновь вызывает запись (3-я рамка), очень сходную с первоначальной. Последняя запись вызванного потенциала на предъявление решетки низкой контрастности после 60 сек экспозиции решетки высокой контрастности не содержит ответа. Отсутствие в записи вызванного потенциала сопровождается попышением субъективного порога (Blakemore and Campbell, 1969).
«В этом исследовании мы намеренно использовали простейший зрительный стимул… Решетка с синусоидально-волновым узором проста потому, что она содержит только одну пространственную частоту, которая представлена в одном меридиане. Наиболее сложным стимулом, исходя из частотного анализа Фурье, является одиночный, резко очерченный круг света, так как он содержит очень широкую полосу пространственных частот и они ориентированы в разных направлениях. Частотная «ширина полосы» индивидуальных пространственных механизмов, выявленная с помощью адаптации, достаточно узка (примерно 1 октава для половины амплитуды). Следовательно, любая сложная световая структура, воздействующая на сетчатку и содержащая широкий спектр компонентов- Фурье, будет активировать многие механизмы. Мы склонны предположить, что структура ответов, получаемых от группы подобных механизмов, может служить для кодирования пространственного содержания определенного сетчаточиого образа и, следовательно, может каким-то неизвестным способом приводить к его идентификации.
Читать дальшеИнтервал:
Закладка:








