К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Перевод с английского Я. Н. Даниловой и Е. Д. Хомской
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Возвращаясь к проприоцептивному механизму периода молчания, следует сказать, что мышечные, веретена прекращают свои разряды во время судороги потому, что они соединены параллельно с главными мышечными волокнами. При сокращении мышцы натяжение веретен уменьшается и они не посылают больше разрядов. Это «параллельное» соединение веретен и является ключом ко всей теории. Важная роль этого механизма состоит в том, что он дает возможность веретенам посылать сигнальную информацию о длине мышц, тогда как если бы веретена были соединены последовательно, они могли бы отвечать только на их напряжение. Таким механизмом, как серворефлекс растяжения, который столь очевидно обеспечивает поддержание определенной длины мышц, а не их напряжения, должны обязательно обладать рецепторы, сигнализирующие о длине мышц» (Merton, 1955, р. 248-259).
Существуют, конечно, и другие рецепторно-эффекторные связи, контролирующие сокращения мышц. Есть рецепторы в сухожилиях и суставах – нельзя не учитывать также и растяжение кожи вокруг мышцы и суставов как источник информации для контроля над движениями. Хотя о, рамма-системе известно больше, чем о других механизмах контроля движений, иннервация, поступающая от мышечных веретен, является, по всей вероятности, основным механизмом, на котором надстраиваются другие контролирующие механизмы (рис. ХII-4).
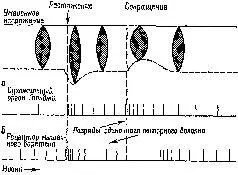
Рис. XII-4. Примеры ответов сенсорных нервных волокон, идущих от сухожильного органа Гольджи (а) и рецептора мышечного веретена (б) и обеспечивающих растяжение и сокращение мышцы. Обратите внимание, что ответ рецептора мышечного веретена (который действует посредством у-системы) выражается как в форме растяжения, так и в форме удлинения мышцы. См. текст на р и с. ХII-5, где обсуждаются эти эффекты (Granit, 1955, см . Thompson, 1967).
Как было установлено, гамма-волокна бывают двух типов. Один тип реагирующих волокон обнаруживает постоянную частоту разрядов, если мышца сохраняет постоянную длину. Другой тип, напротив, обнаруживает уменьшение частоты разрядов во время завершения динамической фазы растяжения. Мэттьюз (1964) в обширном обзоре, посвященном этой теме, приходит к заключению, что такая двойная гамма-иннервация каждого мышечного веретена «обеспечивает относительно независимый контроль за общим режимом работы и отключением сервосистемы», поскольку каждый тип контроля раздельно управляется с помощью центральной стимуляции (рис. ХII-5).
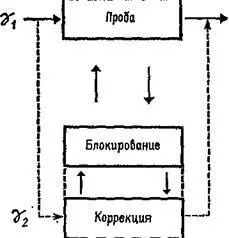
Рис. ХII-5. Двойная иннервация мышечного веретена с позиций концепции Т – О – Т – Е.
Этот анализ важен с двух точек зрения. Во-первых, он привлекает наше внимание к тому факту, что любое, даже самое простое мышечное сокращение должно управляться набором сигналов, которые адресуются либо только к мышечным рецепторам, либо идут одновременно с теми сигналами, которые поступают к сократительным элементам мышцы. Факты (см. следующие разделы) свидетельствуют о том, что некоторые области мозга, а именно базальные ганглии и передняя часть мозжечка, функционируют совместно, непосредственно управляя сократительной альфа-системой и опосредованно воздействуя на рецепторы, контролирующие гамма-разряды, благодаря чему и достигается подобная согласованность движений.
Во-вторых, управление рефлекторным сервомеханизмом легко представить себе, поскольку этот процесс разыгрывается на уровне рецепторов. Следовательно, нет необходимости в специальном устройстве типа клавиатуры фортепиано, с помощью которого импульсы, идущие от мозга, передавали бы мышцам сообщения; дело вовсе не в том, чтобы осуществить сначала одно, а потом другое сокращение, изменить длину того или иного из мышечных волокон. Существующее состояние сокращения мышечного волокна предопределяет вариативное состояние его рецепторов. Набор нервных импульсов взаимодействует с этим фоновым состоянием, изменяя режим работы мышечной сервосистемы и таким образом регулируя мышечное сокращение. Мозг все время должен посылать сигналы, организованные по пространственному и временному принципам, однако сообщения, передаваемые этими сигналами, не должны быть закодированы таким образом, чтобы прямо соответствовать сокращению или расслаблению мышцы, которое эти сообщения вызывают, поскольку эта информация уже содержится в вариативном состоянии рецептора. В следующей главе мы детально проанализируем эти сообщения, которые должны содержать подобный код.
Подведем итоги. Нейрофизиологические данные свидетельствуют о существовании прямых анатомических афферентных путей, идущих от спинного мозга к мышечным рецепторам. Наиболее фундаментальным фактом является то, что основным принципом организации даже самого простого рефлекса является не дуга: стимул – центральная нервная система – реакция, а замкнутая сервосистема в виде последовательности: проба (test) – операция (operate)-проба (test)-результат (exit) (Т-О-Т-Е). Когда с помощью центральной нервной системы рефлексы объединяются в Сложные движения, эта интеграция не может быть эффективной при посылке серий сигналов непосредственно или исключительно сокращающимся мышцам, вызывая в них мелодии, как на клавиатуре фортепиано. Подобные сигналы только бы нарушали сервопроцессы. Чтобы предотвратить эти нарушения, наборы сигналов должны передаваться либо только мышечным рецепторам, либо одновременно с другими сигналами адресоваться непосредственно к мышечным волокнам. Таким образом, интегрированное движение в основном зависит от режима работы, настройки мышечных рецепторов.
Теория, излагаемая в этой книге, вполне согласуется с взглядами Ф. Меттлера (1967). Последний считает, во-первых, что «тонический фон движения служит своего рода канвой… на которой разыгрывается само движение». Этот фон находится под влиянием системы афферентных импульсов, которые отражают «потенциально доминирующие» мозговые процессы. Эта «канва», это фоновое состояние формируется благодаря работе ряда периферических сервомеханизмов, контролирующих мышечные сокращения.
Далее, Меттлер признает существование «ассоциированных массивных движений (предмет данного раздела), на фоне которых вырисовываются отдельные детали…». Массивные процессы регуляции мышечного тонуса осуществляются базальными ганглиями, самыми передними структурами ствола мозга. Наконец, «на этом фоне статических и глобально действующих сил кора мозга ответственна за точность и интенсивность движений, з также за изменение скорости уже начавшегося движения и той скорости, с которой это движение усиливается и прекращается».
Читать дальшеИнтервал:
Закладка:







