К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Перевод с английского Я. Н. Даниловой и Е. Д. Хомской
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
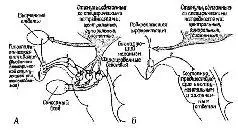
Рис. XIV-8. А – подкрепление моторных структур. Схема взаимосвязей между «подкрепляющей системой мозга» и системой ответов, основанная, на результатах экспериментов; В – роль сенсорного входа. Схема иллюстрирует взаимоотношение между специфическими потребностями и «непосредственным подкреплением», получаемым через сенсорный вход (Valenstein, 1970).
Это предположение родилось в результате серии изобретательных экспериментов, выполненных Э. Валенстайном, которые, однако, в свою очередь поставили ряд новых вопросов. В этих экспериментах электроды вживлялись в ту область гипоталамуса крыс, электрическое раздражение которой вызывает движения еды, питья и попыток грызть кусок дерева. Когда один из таких поведенческих актов наблюдался особенно явно и стабильно, внешние условия эксперимента менялись таким образом, что первоначальный результат нельзя было получить. Так, в опыте создавались такие условия, когда в течение целой ночи животное через нерегулярные интервалы подвергалось стимуляции. На следующее утро та же стимуляция ранее раздражаемого участка вызывала либо первоначальное, либо измененное поведение, что зависело от внешних условий, и, когда были испробованы все возможные варианты, обнаружилось, что часто доминировали новые выработанные движения. Валенстайн пишет:
«В стандартных тестах на выявление обусловленного стимулами поведения удалялся объект (цель, к которой стремилась крыса), и животное-переводилось на ночной режим, когда ему предъявлялись два других целевых объекта. Если, например, крыса во время стимуляции в первой серии опытов пила, то в ночное время сосуд с водой удалялся и в клетке сохранялись только кусочки пищи и дерева. Параметры стимула не менялись. Если животное не проявляло нового поведения, оно переводилось во второй, третий и т. д. раз на ночной режим. Однако в большинстве случаев-одного перевода на ночные условия было достаточно, чтобы возникало новое поведение. После появления нового поведения животным предъявлялись два стандартных теста, в которых начальный целевой объект отсутствовал, и третий стандартный тест (контрольный) со всеми тремя целевыми объектами. В тех случаях, когда животное не подвергалось стимуляции, всегда имелись все три целевых объекта, и, следовательно, животные могли насытиться пищей, напиться воды и удовлетворить свою потребность в том, чтобы грызть кусок дерева, еще до начала опыта. Одиннадцать животных прошли полностью через это испытание, что и составило первый эксперимент…
Вторая серия опытов была проведена после применения различного числа тренировочных стимуляций в условиях отсутствия целевого объекта, к которому прежде стремилось животное… в большинстве случаев вторично сгимулированное поведение появлялось так же закономерно, как и прежнее. Во время предъявления контрольного теста, когда одновременно присутствовали все три целевых объекта, в большинстве случаев с равной? вероятностью наблюдались два типа поведения. Как правило, один тип поведения возникал при действии первых трех-четырех серий стимулов, а затем возникало другое поведение в приблизительно одинаковых по длительности сериях опытов. При предъявлении контрольных тестов животные обнаруживали несколько видов поведения за время предъявления некоторых из 20 стимулов.
Мы хотели выяснить, станет ли доминирующим первый тип поведения, если мы дадим серию контрольных тестов. Хотя в этих опытах наблюдались разнообразные виды поведения, мы пришли к заключению, что как только закреплялся второй вид обусловленного стимулами поведения, печальное поведение, вызванное стимуляцией, переставало быть доминирующим. И, действительно, у ряда животных обнаружилось явное преобладание второго вида поведения во время предъявления контрольных тестов (Valenstein, Cox and Kakolewski, 1969, p. 247-249).
Валенстайн объяснил эти результаты тем, что стимуляция гипоталамуса не вызывает специфических мотивационных состояний типа голода или жажды. Исходя из фактов взаимодействия двух типов явлений – эффекта самораздражения по Олдсу и эффекта вызова с помощью стимуляции определенного вида поведения (еды, питья или попыток грызть дерево), – он приходит к выводу, сходному с тем, который был изложен в гл. X, а именно что электрическая стимуляция, воздействуя на механизмы гомеоста-зиса, создает установку, изменяющую характер ответов.
Обратимся к одному примеру. Мендельсон (1967) описал поведение животных при стимуляции латерального гипоталамуса, которое проявлялось одновременно и в виде самораздражения, и в виде питья. Могенсон и Стивенсон (1967) приводят сходные данные. Мендельсон отобрал животных, у которых реакции питья появлялись при более слабой стимуляции, чем та, которая необходима для поддержания эффекта самораздражения. На этом низком уровне стимуляции животные не нажимали на рычаг, когда не было воды, однако когда жажда утолялась, они также не нажимали на рычаг для получения воды, если только не стимулировался гипоталамус. Мендельсон сделал следующий вывод: «Таким образом, если у крысы вызвать состояние жажды, она будет нажимать на рычаг, чтобы получить воду, а если крысе давать воду, то она будет нажимать на рычаг, чтобы вызвать жажду». Эти результаты очень важны, однако их интерпретация – предположение, что стимуляция вызывает жажду, – не кажется нам достаточно убедительной. Мы могли бы с уверенностью сказать, что у этих животных можно посредством электрической стимуляции вызвать обусловленное стимулом пищевое поведение, и что то же самое поведение можно затем получить и с помощью пищи. Из работ Кунза и Крюса (1968) известно, что животные, проявляющие в ответ на стимулы пищевое поведение, будут подвергать себя самораздражению на уровне ниже «порога вознаграждения», если они получают пищу. Поэтому более разумно было бы предположить, что подкрепление, возникающее результате выполнения вызванного поведенческого акта, обусловливает не голод или жажду, а суммируется с подкреплением, возникающим непосредственно в результате самой стимуляции. Из этого следует, что сумма покрепления от выполнения поведенческих реакций и подпорогового подкрепления, вызванного стимуляцией мозга, может оказаться достаточной, чтобы поддерживать инструментальное поведение. Действительно, ранее Мендельсон (1966) показал, что сытое животное обнаруживает вызванное -стимулами пищевое поведение скорее при комбинации пищи и стимуляции мозга, чем при одной мозговой стимуляции (Valenstein, Cox and Kakolewski, 1969, p. 267-277).
Однако, предполагая, что стимуляция мозга не вызывает состояния жажды, Валенстайн может выбросить вместе с водой и ребенка. У. Роберте (1969) указывает, что сильная электрическая стимуляция может разрушить четко организованное поведение и расширить гипоталамическую зону, при раздражении которой постоянно наблюдаются частные движения, включенные в тот или иной поведенческий акт. Роберте обвиняет Валенстайна в том, что он стоит па позициях Лешли, отрицая специфичность нервных центров. Эта позиция, как он считает, должна быть пересмотрена, потому что, как отметил П. Милнер (Roberts, I960, р. 17-19), она «заставила многих исследователей на несколько десятилетий отказаться от изучения проблемы функциональной специфичности и локализации функций». Вопреки этому Роберте считает, что компетентная ткань состоит из «перекрывающих друг друга, но специфических по своим функциям механизмов».
Читать дальшеИнтервал:
Закладка:







