К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Перевод с английского Я. Н. Даниловой и Е. Д. Хомской
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Эта загадка была частично разрешена, когда было установлено, что результаты разрушения мозга неоднозначны. Локализация функций различных сенсорных модальностей была обнаружена и внутри так называемой ассоциативной коры. Один участок в теменной доле мозга оказывает исключительное влияние на различение кожно-кинестетических сигналов; другой участок в передней части височной доли имеет отношение только к вкусовым ощущениям. Средневисочные участки коры специализированы исключительно на слухе, а участок в нижней части височной доли имеет отношение только к зрению (рис. XVII-5). Более того, повреждения в зтой так называемой ассоциативной коре не вызывают никаких нарушений взаимодействия между раздражителями разных модальностей (Pribram, 1969a).
Таким образом, проблема сводится к определению функций этих специфических сенсорных зон. Экспериментальные исследования для разрешения зтой проблемы проводились в основном на зрительных процессах и поэтому касались коры нижней части височной области. Однако и в отношении слуховых и соматосен-сорных процессов было сделано достаточно для того, чтобы прийти к выводу, что результаты, полученные при изучении зрения, применимы и к другим частям ассоциативной коры, обслуживающим иные сенсорные модальности.
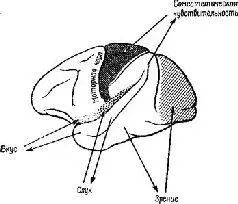
Рис. XVII-5. Схема латеральной поверхности мозга обезьяны, на которой изображены модально-специфические первичные сенсорные зоны и специфические сенсорные подразделения ассоциативной коры.
Прежде всего возник вопрос, вызывают ли повреждения нижневисочной коры мозга обезьян нарушение всех форм зрительных навыков. Скоро удалось установить, что такое повреждение не нарушало слежения за зрительным объектом, например за летящим комаром. Позднее это было подтверждено посредством фотографирования отражения наблюдаемых объектов от роговицы (Bagshaw, Mackworth and Pribram, 1970).
Нарушения возникали лишь в том случае, когда перед обезьянами с повреждениями мозга стояла задача выбора или дифференцировки. Такие нарушения были обнаружены при самых различных формах зрительного выбора независимо от того относился ли он к цвету, форме, контуру или яркости (Mishkin and Pribram, 1954). Единственным общим в этих задачах было то, что все они были зрительными и во всех предусматривался выбор, а также то, что массивность нарушения поведения была пропорциональна трудности, которую испытывали при обучении нормальные обезьяны.
Эти результаты сразу же заставили исследователей предположить, что затруднения, наблюдаемые у обезьян с повреждениями мозга, выражаются скорее в их неспособности заучить (и запомнить) задачу, чем в неспособности воспринять основные ключевые признаки, которые ведут к ее разрешению. Для проверки этой гипотезы было проведено исследование того, как обезьяны с удалением нижневисочной коры и с частичным удалением первичной зрительной зоны решают различные зрительные задачи. В общем результаты подтвердили гипотезу, что научение и восприятие могут быть разграничены путем повреждений разных частей коры. Так, повреждение первичной зрительной коры сказывалось на решении задач на восприятие: обезьяна не могла подтянуть орех, привязанный к шнурку, находящемуся среди нескольких других, к которым не была привязана приманка; разрушение нижневисочных отделов коры не вызывало таких нарушений. Противоположная картина наблюдалась при выполнении заданий на научение (Wilson, Mishkin, 1959, рис. XVII-6).
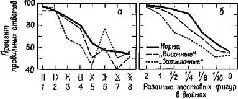
Рис. XVII- 6. A – средние результаты выполнения структурированного серийного теста у трех групп обезьян (контрольная группа; обезьяны с поражением первичной коры; обезьяны с поражением нижновисочной зрительной коры); В – средние результаты выполнения задания на различение размера у трех групп обезьян (Wilson, Mishkin, 1959).
Следует, однако, обратить внимание на то, что здесь сказано «в общем». Расхождение между влияниями, оказываемыми этими двумя типами повреждений на восприятие и научение, не было ни полным, ни вполне определенным. Результатом повреждения первичной коры было некоторое замедление процесса научения, что можно было ожидать, так как обезьяны приспосабливались к сниженной остроте зрения и большим скотомам в зрительном поле. Более загадочным было то, что у обезьян с повреждениями нижневисочной коры наблюдалось некоторое снижение остроты зрения, например в отношении размера (Mishkin, Hall, 1955) н критической частоты мельканий (Mishkin and Weiskrantz, 1959). Поскольку острота зрения проверялась методом выработки дифференцировок, эта загадка легко объяснялась тем, что был нарушен механизм выбора, а в результате этого нарушалось как научение, так и запоминание. Однако проверка на людях, у которых по той или иной причине была удалена височная доля, поставила под сомнение такое объяснение (Goldman et al., 1968), Изменения критической частоты мельканий наблюдались, даже когда использовался словесный отчет о слиянии (метод определения порогов при повышении и снижении частоты мельканий). Читателя не должны слишком удивлять эти результаты, как не должны были они удивлять нас, исследователей. Распознавание образов, дифференцировка знаков требуют участия как процесса формирования Образа, так и памяти. Знакомое лицо стало знакомым благодаря опыту, но оно воспринимается целостно и непосредственно.
Мы были озадачены тем, что вначале не могли представить себе механизм, который частично – но не полностью – разделял бы восприятие и научение. В классическом понимании функций коры вводимая информация воспринимается посредством первичной коры; к тому, что уже заучено, добавляется затем сумма других видов информации и ассоциаций, хранимых в ассоциативной коре. У. Пенфилд (1969) как-то назвал кору височной доли «корой памяти», так как электрическая стимуляция этой части мозга у эпилептиков вызывает воспроизведение картин пережитого. Однако эта классическая точка зрения не согласуется с парадоксальными фактами, обнаруженными в ходе экспериментов с рассечением коры.
Как указывалось во введении к этой главе, можно полностью разрешить этот парадокс, если серьезно рассмотреть гипотезу о том, что влияние нижневисочной коры на зрительные процессы обусловлено не столько тем, что она получает зрительную информацию от первичной коры, сколько тем, что она воздействует через кортикофугальные связи на процессы переработки зрительной информации, происходящие в подкорковых структурах. Какие же факты делают вероятным существование такого коркового контроля подкорковых механизмов зрения? И как можно представить себе его функционирование на основании уже известных нейропсихологических данных?
Читать дальшеИнтервал:
Закладка:







