Николай Курчанов - Генетика человека с основами общей генетики. Учебное пособие

- Название:Генетика человека с основами общей генетики. Учебное пособие
- Автор:
- Жанр:
- Издательство:Литагент «СпецЛит»d5a9e1b1-0065-11e5-a17c-0025905a0812
- Год:2009
- Город:Санкт-Петербург
- ISBN:978-5-299-00411-3
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Николай Курчанов - Генетика человека с основами общей генетики. Учебное пособие краткое содержание
В пособии освещаются все разделы современной генетики, необходимые для понимания генетики человека и психогенетики. Показана методологическая роль генетики в современной биологии. Первые главы посвящены фундаментальным положениям общей генетики. В специальных разделах рассматриваются вопросы медицинской генетики, генной инженерии, генетики поведения, эволюции, психогенетики.
Второе издание книги значительно переработано автором с учетом новой информации, опубликованной за последние три года.
Пособие предназначено для студентов биологических, педагогических, психологических и социологических факультетов. Представляет интерес для научных работников всех специальностей, занимающихся вопросами, связанными с изучением биологической природы человека.
2-е издание, переработанное и дополненное.
Генетика человека с основами общей генетики. Учебное пособие - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
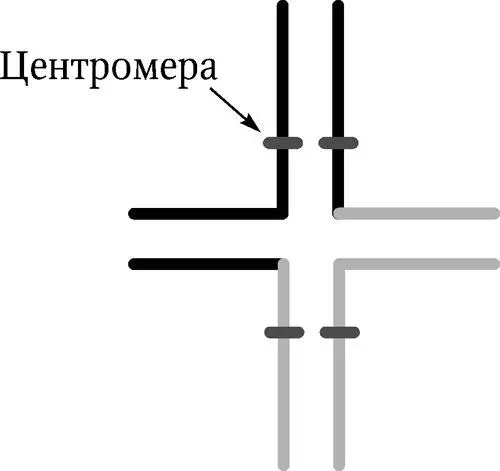
Рис. 5.3. Синапсис хромосом при реципрокной транслокации в профазе мейоза. На каждой хромосоме отмечена центромера
Гетерозиготы по реципрокным транслокациям обладают пониженной плодовитостью, так как продуцируют дефектные гаметы. Только у двух видов гамет из шести возможных при разных способах расхождения хромосом имеются полные комплекты генов. Остальные несут дупликации и нехватки и не могут дать жизнеспособного потомства. У животных реципрокные транслокации встречаются редко, обычно с негативным эффектом, но они широко распространены у растений. Благодаря специальным механизмам, транслокация обеспечивает изоляцию новых форм.
Участок хромосомы может менять свое положение и без реципрокного (взаимного) обмена, оставаясь на той же хромосоме или включаясь в другую. Такое явление называется транспозицией.Транспозиции будут рассматриваться ниже как важный самостоятельный раздел современной генетики.
Вероятно, все типы хромосомных перестроек имеют единый механизм и обусловлены лабильностью генома.
Причиной изменения фенотипа при различных хромосомных перестройках часто является изменение расположения гена. Этот феномен получил название эффект положения гена. Он показан для многих генов и обычно влияет на их регуляторную систему. Например, при перемещении гена из эухроматина в гетерохроматиновую область его активность утрачивается, хотя сам ген не изменяется.
Геномные мутации –это изменения числа хромосом. Среди геномных мутаций также выделяют несколько разновидностей (рис. 5.4).
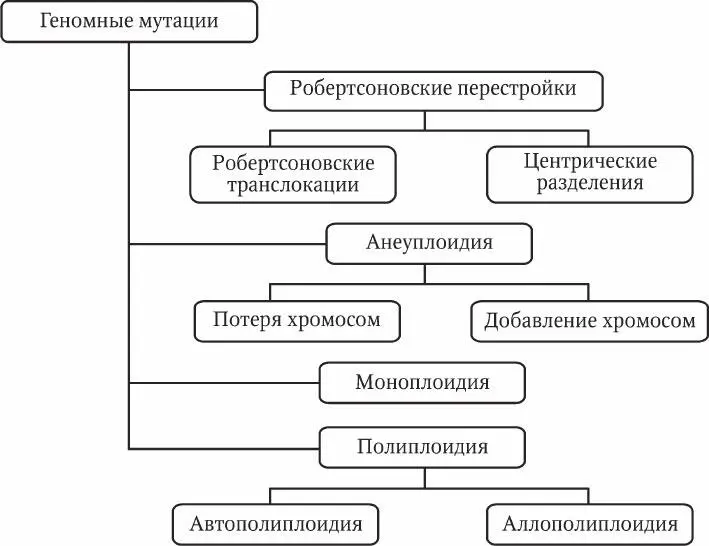
Рис. 5.4. Геномные мутации
Робертсоновские перестройки – слияния и разделения хромосом в области центромеры. Названы они по имени В. Робертсона, который предложил свою гипотезу механизма таких мутаций. Центрические слияния ( робертсоновские транслокации ) представляют собой слияния двух негомологичных акроцентрических хромосом с образованием одной субметацентрической хромосомы. При разделении, наоборот, одна субметацентрическая хромосома делится на две акроцентрические хромосомы. При этом должна образоваться новая центромера, иначе хромосома без центромеры будет потеряна при митозе.
Робертсоновские перестройки приводят к изменению числа хромосом в кариотипе, не влияя на общее количество генетического материала в клетке. Оба варианта перестроек представлены в природе, но робертсоновские транслокации встречаются значительно чаще. Они являются одним из магистральных путей эволюции кариотипов.
Анеуплоидия – изменение числа хромосом, не кратное гаплоидному набору. Как правило, представляет собой добавление или потерю одной или двух хромосом диплоидного набора. У животных анеуплоидия обычно приводит к тяжелым аномалиям или летальности. Однако у растений трисомия (наличие трех гомологичных хромосом) может служить фактором генетического разнообразия. Причиной анеуплоидии является нерасхождение хромосом в мейозе и образование несбалансированных гамет.
Моноплоидия (гаплоидия) – гаплоидное состояние диплоидного организма. Эта мутация интенсивно изучается у растений, так как позволяет видеть проявление рецессивных аллелей. У животных моноплоидия обычно приводит к летальному исходу.
Автополиплоидия – наличие в клетке более двух одинаковых гаплоидных наборов. Эта разновидность мутации довольно широко представлена в природе у протистов, грибов и растений. Плоидность макронуклеуса инфузорий может достигать нескольких сотен. У животных встречается редко и обычно приводит к летальному исходу на ранних стадиях эмбриогенеза. У культурных растений сбалансированные полиплоиды (т. е. кариотипы с четным числом гаплоидных наборов – 4n, 6n, 8n и т. п.) получают искусственным путем из-за их более крупных размеров. Несбалансированные полиплоиды (3n, 5n, 7n и т. п.) растений часто имеют пониженную фертильность вследствие нарушений мейоза. Тем не менее некоторые растения-триплоиды (3n) обладают большими размерами и продуктивностью по сравнению с диплоидными (2n) и тетраплоидными (4n).
Аллополиплоидия – объединение в клетке разных геномов посредством гибридизации. В природе для многих цветковых растений описаны полиплоидные ряды различной степени плоидности. Эти ряды возникают путем гибридизации разных видов и последующего удвоения родительских гаплоидных наборов. Так преодолевается барьер бесплодия при скрещивании разных видов.
Цитоплазматические мутации– это изменения неядерных генов. Генетическая информация органелл (в отличие от хромосомной) представлена в зиготе в разном объеме от каждого из родителей. Чаще всего преобладает вклад материнского организма, но у некоторых видов наследуются внеядерные геномы от обоих родителей. Мутации генома органелл обычно представлены точечными мутациями и делециями. Основными носителями внеядерной генетической информации в клетках высших эукариот являются хлоропласты и митохондрии.
В хлоропластах фенотипическое проявление мутаций выражается нарушением фотосинтеза, изменением чувствительности к температуре и устойчивости к антибиотикам. Генетика хлоропластов разработана хуже, чем генетика митохондрий.
У позвоночных митохондрии служат единственным хранилищем внеядерного генетического материала. Источником митохондрий в зиготе, вероятно, служит только яйцеклетка. Хотя 95 % митохондриальных белков кодируется в ядре, мутации мт-ДНК происходят в 10 раз чаще, чем в ядерных генах, поэтому их вклад ощутим.
Фенотипическое проявление мутаций мт-ДНК во многом зависит от уровня потребления АТФ той или иной тканью. Поскольку наиболее энергозависимыми являются нервная и мышечная системы, такие мутации наиболее часто проявляются в форме различных нейропатий и миопатий.
5.2. Рекомбинация
Рекомбинация генетического материала в природе происходит на разных уровнях.
Рекомбинация несцепленных геновпроисходит благодаря механизму независимого распределения хромосом в мейозе и формирования новых генотипов при оплодотворении. Даже единичные мутантные аллели в популяции резко увеличивают генотипическое разнообразие. Именно этот фактор и является главным эволюционным преимуществом полового размножения.
Замечено, что чем выше уровень организации организмов, тем большее значение приобретает рекомбинация. Если у прокариот жизненно важные функции могут определяться единичными генами, то у высших животных фенотипические признаки обычно определяются сложными сочетаниями генов. Некоторые новые сочетания генов могут превысить предковый генотип по адаптационной ценности. Как сказал известный эволюционист В. Грант, «рекомбинация – это механизм, который производит сборку генных сочетаний » (Грант В., 1980).
Читать дальшеИнтервал:
Закладка:









