Феликс Филатов - КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле.

- Название:КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле.
- Автор:
- Жанр:
- Издательство:ОАО ордена Знак почета Смоленская областная типография им. В.И.Смирнова
- Год:2011
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Феликс Филатов - КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле. краткое содержание
КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле. - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
После транскрибирования мРНК не остается комплементарно связанной с ДНК-шаблоном; она освобождается от ДНК, которая затем восстанавливает свою двойную структуру. В одной эукаритической клетке количество молекул мРНКможет составлять свыше 10.000. Наряду с молекулами мРНК на ДНК образуются и другие транскрипты, в том числе молекулы рибосомной и транспортных РНК, которые также имеют важное значение в реализации генетической информации. Все эти РНК называют еще ядерными. Наиболее обильными РНК в клетках всех видов являются молекулы рибосомной РНК (рРНК), которые выполняют роль структурных компонентов рибосом. У эукариот синтез рРНК контролируется огромным количеством генов (сотни и тысячи копий) и происходит в ядрышке. Не похоже, что структура рРНК имеет серьезное значение для обсуждаемой далее формальной организации генетического кодирования, и мы не станем на ней останавливаться. Совершенно другую роль в этих процессах имеют встречающиеся в клетке в несколько меньших количествах молекулы транспортных РНК (тРНК), которые участвуют в декодировании информации, трансляции. Это те самые декодеры или молекулы-посредники ( codemakers Барбьери), — которые обусловливают специфическую связь между хранилищем генетической информации, нуклеиновыми кислотами, и строительным материалом, служащим для ее реализации, то есть для конструирования пептидов — аминокислотами.
Роль транспортных РНК в синтезе белка была постулирована еще до их открытия. В 1955г. Крик приписал им функцию адаптера, который может нести аминокислоту и образовывать водородные связи с кодирующей полинуклеотидной матрицей. Гипотеза адаптера оказалась необходимой в связи с упоминавшейся уже невозможностью обнаружить между аминокислотами и нуклеиновыми кислотами стереохимическое соответствие, достаточное для того, чтобы обеспечить считывание генетического кода . В 1957г. в лаборатории МэлонаХогланда было показано, что в ходе белкового синтеза активированные аминокислоты переносятся на особый тип РНК, получивший тогда наименование растворимой РНК и называемой теперь транспортной.
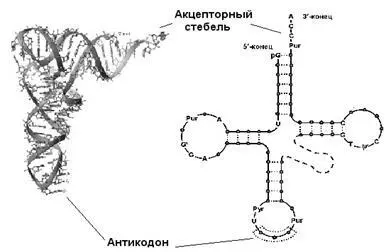
Стереохимия тРНК хорошо изучена и весьма характерна. Мы остановимся на ней поподробнее. Типичная молекула тРНК — это полинуклеотидная цепь длиной 75-90(по преимуществу, 76) нуклеотидов. Молекулярные массы тРНК лежат в пределах 17.000-35.000.Часть оснований нуклеотидных пар, уже после синтеза тРНК, в определенных положениях модифицирована, это неканонические, редкие, так называемые ( минорные ), составляющие до 10% от общего числа. Среди них — дигидроуридин ( D ), псевдоуридин ( Ψ ) и инозин ( I ); последний играет существенную роль в узнавании кодона. В дополнение к этим модификациям несколько нуклеозидов метилированы. Все эти модификации — результат посттранскрипционного процессинга тРНК, которая копируется с «нормальной» матрицы. В 75% случаев молекулы тРНК открываются5′-гуанином (он фосфорилирован) и во всех случаях завершается триплетом ССА -3′.
Вторичная структура этой молекулы сформирована четырьмя короткими двуцепочечными стеблями и напоминает клеверный лист.
Каждый из четырех стеблей состоит из 4-7уотсон-криковских пар, образующих двойные спирали. Сами стебли носят названия акцепторного, антикодонного, а также D(содержащий дигидроуридин) и T (содержащий риботимидин). Некоторые нуклеотиды консервативны и их позиции в составе тРНК остаются инвариантными — либо полуинвариантными, если сохраняется их пуриновая или пиримидиновая природа. На акцепторном стебле тРНК имеется участок связывания с аминокислотой; он неспецифичен и для всех аминокислот один и тот же: ССА -3′. Противоположный стебель содержит одноцепочечную петлю с антикодоном, распознающим кодон на мРНК. Две другие, боковые, петли предназначены для связывания с рибосомой и с аминоацил-тРНКсинтетазой (АРСазой). Четвертая, не всегда выраженная, петля так и называется — дополнительная или вариабельная (V). У тРНК, узнаваемых АРСазами класса I, она, как правило, короче (4-5 нуклеотидов),у тРНК, узнаваемых АРСазами класса II — длиннее (13-21 нуклеотидов).
Третичная (пространственная) структура любой тРНК складывает все ее четыре ветви (стебли с петлями) в так называемую Г-форму (L-форму,если использовать латиницу):
Г-форма состоит из двух почти перпендикулярных друг другу спиралей А-РНК (11 пар оснований на виток). Два конца буквы Г образованы ССА -3`-концом и антикодонной петлей, которые находятся на расстоянии 80Å друг от друга. Наружный край угла буквы Г образован Т-петлей. Акцепторный и Т-стебли уложены один вслед за другим и образуют единую двойную спираль. В примерно такую же структуру (только с расхождением осей на 26°) уложены антикодонный и D-стебли.Эта структура на предыдущем рисунке обозначена жирной черной кривой.
Уже цитированный Др.Зенгер назвал тРНК « сокровищницей стереохимической информации ». Он отмечает, что кроме уотсон-криковских пар, ответственных за большую часть горизонтальных взаимодействий между основаниями (особенно в стеблях), в тРНК имеется ряд нестандартных пар и триплетов . Такие пары располагаются в основном с наружной стороны угла и в шарнирной области буквы Г. «Качающаяся пара» G 4— U 69дрожжевой фенилаланиновойтРНК входит в состав акцепторного стебля. По структуре она сходна с уотсон-криковской и не нарушает двойной спирали, а только создает небольшую выпуклость в сахарофосфатном остове. Обратная непланарная — хугстеновская — пара m 1 A 58- T 54возникает благодаря блокаде уотсон-криковской из-за метилированного аденина-58, и хугстеновское спаривание оказывается единственно возможным способом образования водородных связей с другим основанием (курсив здесь далее в этой главе — до новой ссылки — цитата из книги Зенгера). Пурин-пуриновая пара m 2 G 26- A 44 "длиннее«, чем обычная уотсон-криковская и в значительной мере непланарна; именно она ответственна за 26°-ое расхождение антикодонового и D-стеблей.
В тРНК имеется несколько триплетов, в которых уотсон-криковскую пару дополняет третье основание, присоединенное со стороны главного желобка либо одной ( m 2 G 10-С 25- G 45 ), либо двумя ( G 22-С 13- m 7 G 46 и A 23- U 12- A 9 ) водородными связями;во всех этих триплетах представлена и параллельная, и антипараллельная ориентация полинуклеотидных цепей. Большинство «третичных» водородных связей образуется между консервативными основаниями.
Многочисленные вертикальныестэкинг-взаимодействия, наличие которых чрезвычайно характерно для архитектуры тРНК, укрепляют эту структуру. В дрожжевой фенилаланиновойтРНК только 42 основания из 76 входят в спиральные участки, но в стэкинг-взаимодействии участвует 71 основание. Схема стэкинга в спиральных доменах тРНК практически такая же, как и у двуцепочечных А-РНК, однако, перекрывание оснований несколько сильнее, и плотность спирали эквивалентна структуре с числом пар, несколько меньшим, чем 11 . В отдельных местах молекулы тРНК основание одной цепи встраивается между двумя основаниями другой (интеркаляция), при этом возникает особый вид стэкинга.
Читать дальшеИнтервал:
Закладка:





